Variations in the Phytoremediation Efficiency of Metal-polluted Water with Salvinia biloba: Prospects and Toxicological Impacts

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plants Collection and Characterization
2.2. Metal Stock Solution Preparation
2.3. Metal Removal Studies
2.4. Metal Removal Kinetics
2.5. Elemental Analysis
2.6. Physiological Parameters
2.6.1. Quantification of Photosynthetic Pigments
2.6.2. Determination of Antioxidants Pigments
2.6.3. Soluble Carbohydrates
2.7. Statistical Analysis
3. Results and Discussion
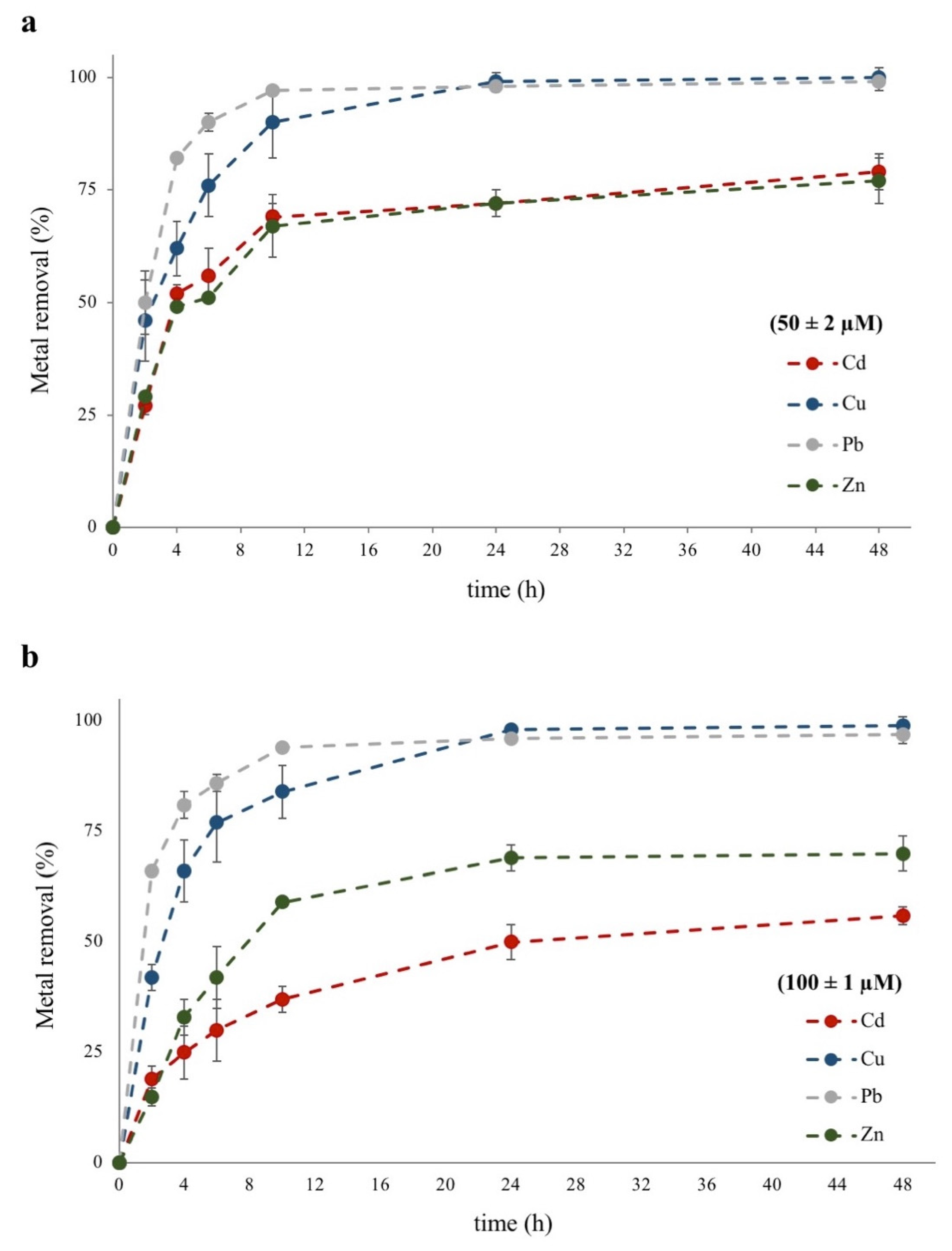
3.1. Metal Removal Efficiency from Water-Polluted Samples
3.2. Metal Removal Kinetics
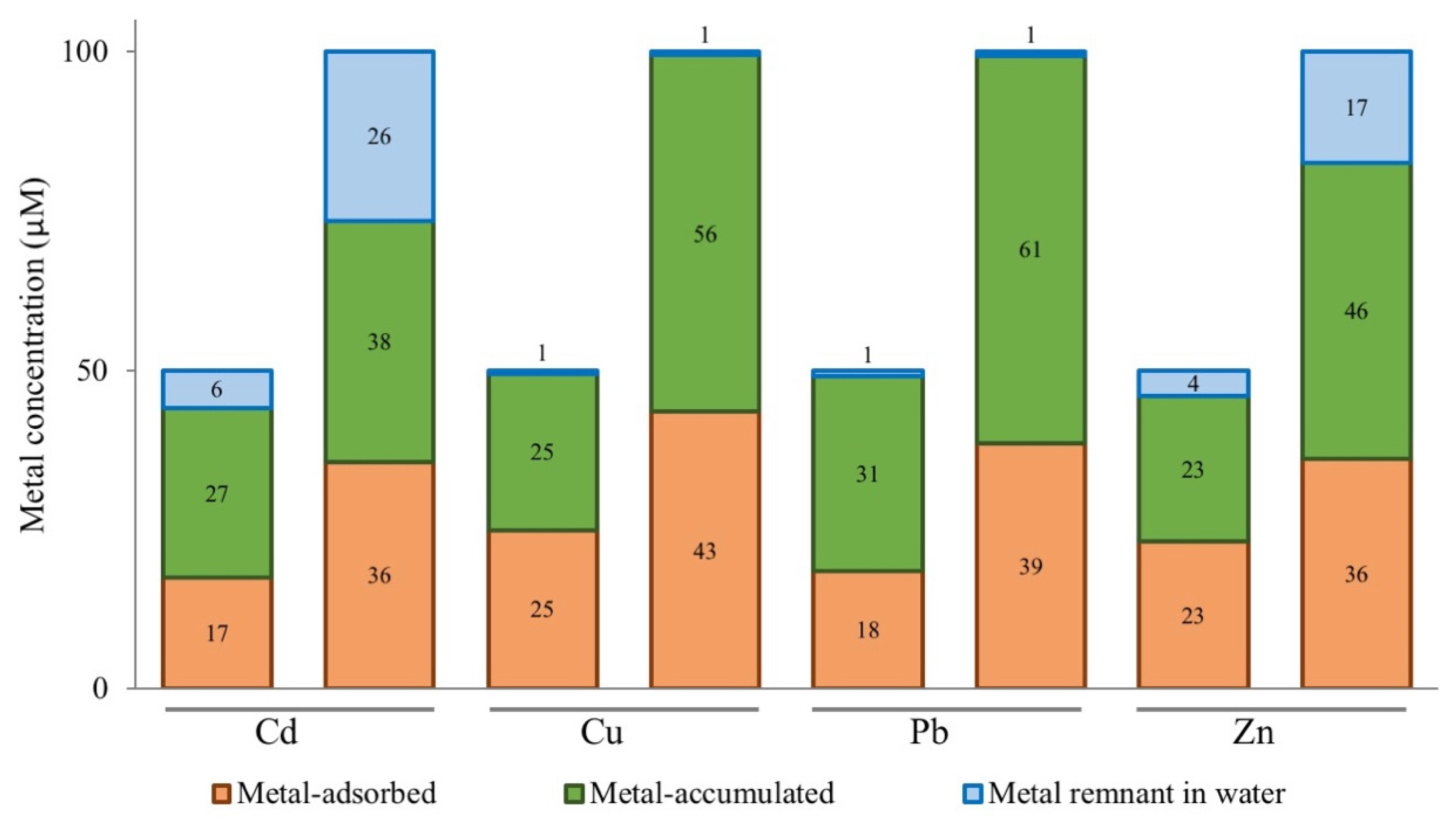
3.3. Elemental Analysis
3.4. Evaluation of Metal Phytotoxicity in S. Biloba
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jaishankar, M.; Tseten, T.; Anbalagan, N.; Mathew, B.B.; Beeregowda, K.N. Toxicity, mechanism and health effects of some heavy metals. Interdiscip. Toxicol. 2014, 7, 60–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azimi, A.; Azari, A.; Rezakazemi, M.; Ansarpour, M. Removal of heavy metals from industrial wastewaters: A review. ChemBioEng Rev. 2017, 4, 37–59. [Google Scholar] [CrossRef]
- Ali, H.; Khan, E.; Sajad, M.A. Phytoremediation of heavy metals—Concepts and applications. Chemosphere 2013, 91, 869–881. [Google Scholar] [CrossRef] [PubMed]
- Barakat, M.A. New trends in removing heavy metals from industrial wastewater. Arab. J. Chem. 2011, 4, 361–377. [Google Scholar] [CrossRef] [Green Version]
- Sukumaran, D. Phytoremediation of heavy metals from industrial effluent using constructed wetland technology. Appl. Ecol. Environ. Sci. 2013, 1, 92–97. [Google Scholar] [CrossRef]
- Sharma, P.; Pandey, S. Status of phytoremediation in world scenario. Int. J. Environ. Biorem. Biodeg. 2014, 2, 178–191. [Google Scholar] [CrossRef]
- Dhir, B. Potential of biological materials for removing heavy metals from wastewater. Environ. Sci. Pollut. Res. Int. 2014, 21, 1614–1627. [Google Scholar] [CrossRef]
- Dixit, R.; Wasiullah, E.; Malaviya, D.; Pandiyan, K.; Singh, U.B.; Sahu, A.; Shukla, R.; Singh, B.P.; Rai, J.P.; Sharma, P.K.; et al. Bioremediation of heavy metals from soil and aquatic environment: An overview of principles and criteria of fundamental processes. Sustainability 2015, 7, 2189–2212. [Google Scholar] [CrossRef] [Green Version]
- Rai, P.K. Heavy metal phytoremediation from aquatic ecosystems with special reference to macrophytes. Crit. Rev. Environ. Sci. Technol. 2009, 39, 697–753. [Google Scholar] [CrossRef]
- Marchetti, Z.Y.; Latrubesse, E.M.; Pereira, M.S.; Ramonell, C.G. Vegetation and its relationship with geomorphologic units in the Parana River floodplain, Argentina. J. S. Am. Earth Sci. 2013, 46, 122–136. [Google Scholar] [CrossRef]
- Dhir, B. Salvinia: An aquatic fern with potential use in phytoremediation. Environ. We Int. J. Sci. Technol. 2009, 4, 23–27. [Google Scholar]
- Maine, M.A.; Duarte, M.; Suñe, N.L. Cadmium uptake by floating macrophytes. Water Res. 2001, 35, 2629–2634. [Google Scholar] [CrossRef]
- Maine, M.A.; Suñe, N.L.; Lagger, S.C. Chromium bioaccumulation: Comparison of the capacity of two floating aquatic macrophytes. Water Res. 2004, 38, 1494–1501. [Google Scholar] [CrossRef] [PubMed]
- Paris, C.; Hadad, H.R.; Maine, M.A.; Suñe, A.N. Eficiencia de dos macrófitas flotantes libres en la absorción de metales pesados. Limnetica 2005, 24, 237–244. [Google Scholar] [CrossRef]
- Suñe, N.L.; Sánchez Galván, G.; Caffaratti, S.; Maine, M.A. Cadmium and chromium removal kinetics from solution by two aquatic macrophytes. Environ. Pollut. 2007, 145, 467–473. [Google Scholar] [CrossRef]
- Hadad, H.R.; Maine, M.A.; Natale, G.S.; Bonetto, C. The effect of nutrient addition on metal tolerance in Salvinia herzogii. Ecol. Eng. 2007, 31, 122–131. [Google Scholar] [CrossRef]
- Freitas, F.; Lunardi, S.; Souza, L.B.; von der Osten, J.S.C.; Arruda, R.; Andrade, R.L.T.; Battirola, L.D. Accumulation of copper by the aquatic macrophyte Salvinia biloba Raddi (Salviniaceae). Braz. J. Biol. 2018, 78, 133–139. [Google Scholar] [CrossRef] [Green Version]
- Tello Zevallos, W.; Salvatierra, L.M.; Loureiro, D.B.; Morató, J.; Pérez, L.M. Evaluation of the autochthonous free-floating macrophyte Salvinia biloba Raddi for use in the phytoremediation of water contaminated. Desalin. Water Treat. 2018, 103, 282–289. [Google Scholar] [CrossRef] [Green Version]
- Castillo Loría, K.; Emiliani, J.; Herrero, M.S.; Bergara, C.D.; Salvatierra, L.M.; Pérez, L.M. Effect of daily exposure to Pb-contaminated water into Salvinia biloba physiology and phytoremediation performance. Aquatic Toxicol. 2019, 210, 158–166. [Google Scholar] [CrossRef]
- Liu, Z.-R.; Chen, X.-S.; Zhou, L.-M.; Wei, P. Development of a first-order kinetics-based model for the adsorption of nickel onto peat. Min. Sci. Technol. 2009, 19, 230–234. [Google Scholar] [CrossRef]
- Zheng, X.; Bi, C.; Li, Z.; Podariu, M.; Hage, D.S. Analytical methods for kinetic studies of biological interactions: A review. J. Pharm. Biomed. Anal. 2015, 113, 163–180. [Google Scholar] [CrossRef] [Green Version]
- Wintermans, J.F.; De Mots, A. Spectrophotometric characteristics of chlorophyll a and b and their phenophytins in ethanol. Biochim. Biophys. Acta 1965, 109, 448–455. [Google Scholar] [CrossRef]
- Bizzo, A.L.; Intorne, A.C.; Gomes, P.H.; Susuki, M.S.; Esteves, B.D. Short-term physiological responses to copper stress in Salvinia auriculata Aubl. Acta Limnol. Bras. 2014, 26, 268–277. [Google Scholar] [CrossRef] [Green Version]
- Casati, P.; Walbot, V. Differential accumulation of maysin and rhamnosylisoorientin in leaves of high-altitude landraces of maize after UV-B exposure. Plant Cell Environ. 2005, 28, 788–799. [Google Scholar] [CrossRef]
- Kratochvil, D.; Volesky, B. Advances in the biosorption of heavy metals. Trends Biotechnol. 1998, 16, 291–300. [Google Scholar] [CrossRef]
- Sánchez-Galván, G.; Monroy, O.; Gómez, J.; Olguín, E.J. Assessment of the hyperaccumulating lead capacity of Salvinia minima using bioadsorption and intracellular accumulation factors. Water Air Soil Pollut. 2008, 194, 77–90. [Google Scholar] [CrossRef]
- Olguín, E.J.; Sánchez-Galván, G.; Pérez-Pérez, T.; Pérez-Orozco, A. Surface adsorption, intracellular accumulation and compartmentalization of lead in batch-operated lagoons with Salvinia minima as affected by environmental conditions, EDTA and nutrients. J. Ind. Microbiol. Biotechnol. 2005, 32, 577–586. [Google Scholar] [CrossRef] [PubMed]
- Estrella-Gómez, N.; Mendoza-Cózatl, D.; Romero-Sánchez, R.; Gonzáles-Mendoza, D.; Zapata-Pérez, O.; Martínez-Hernández, A.; Santamaría, J.M. The Pb-hyperaccumulator aquatic fern Salvinia minima Baker, responds to Pb2+ by increasing phytochelatins via changes in SmPCS expression and in phytochelatin synthase activity. Aquatic Toxicol. 2009, 91, 320–328. [Google Scholar] [CrossRef]
- Leal-Alvarado, D.A.; Estrella-Maldonado, H.; Sáenz-Carbonell, L.; Ramírez-Prado, J.H.; Zapata-Pérez, O.; Santamaría, J.M. Genes coding for transporters showed a rapid and sharp increase in their expression in response to lead, in the aquatic fern (Salvinia minima Baker). Ecotoxicol. Environ. Saf. 2018, 147, 1056–1064. [Google Scholar] [CrossRef]
- Calvo, J.; Jung, H.; Meloni, G. Copper metallothioneins. IUBMB Life 2017, 69, 236–245. [Google Scholar] [CrossRef] [Green Version]
- Schor-Fumbarov, T.; Goldsbrough, P.; Adam, Z.; Tel-Or, E. Characterization and expression of a metallothionein gene in the aquatic fern Azolla filiculoides under heavy metal stress. Planta 2005, 223, 69–76. [Google Scholar] [CrossRef]
- Phetsombat, S.; Kruatrachue, M.; Pokethitiyook, P.; Upatham, S. Toxicity and bioaccumulation of cadmium and lead in Salvinia cucullata. J. Environ. Biol. 2006, 27, 645–652. [Google Scholar] [PubMed]
- Wolff, G.; Pereira, G.C.; Castro, E.M.; Louzada, J.; Coelho, F.F. The use of Salvinia auriculata as a bioindicator in aquatic ecosystems: Biomass and structure dependent on the cadmium concentration. Braz. J. Biol. 2012, 72, 71–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, D.A.; Vasconcelos Filho, S.C.; Rodrigues, A.A.; Rampazzo, D.F.; Rodrigues, C.L.; Vasconcelos, J.M.; Magalhães, P.A.N.R. Effect of cadmium on the morphology and anatomy of Salvinia auriculata. Afr. J. Biotechnol. 2016, 15, 891–896. [Google Scholar] [CrossRef] [Green Version]
- Rsat, H.; Petzold, L.; Pettigrew, M.F. Kinetic modeling of biological systems. Methods Mol. Biol. 2009, 541, 311–335. [Google Scholar] [CrossRef] [Green Version]
- Joseph, L.; Jun, B.M.; Flora, J.R.V.; Park, C.M.; Yoon, Y. Removal of heavy metals from water sources in the developing world using low-cost materials: A review. Chemosphere 2019, 229, 142–159. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Wang, J.; Yu, J.; Wang, Y.; Chi, R. Effects of surface modification on heavy metal adsorption performance and stability of peanut shell and its extracts of cellulose, lignin, and hemicellulose. Environ. Sci. Pollut. Res. 2020. [Google Scholar] [CrossRef] [PubMed]
- Cobbett, C.S. Phytochelatins and their roles in heavy metal detoxification. Plant Physiol. 2000, 123, 825–832. [Google Scholar] [CrossRef] [Green Version]
- Mendoza-Cózatl, D.; Rodríguez-Zavala, J.; Rodríguez-Enríquez, S.; Mendoza-Hernández, G.; Briones-Gallardo, G.; Moreno-Sánchez, R. Phytochelatin-cadmium-sulfide high-molecular-mass complexes of Euglena gracilis. FEBS J. 2006, 273, 5703–5713. [Google Scholar] [CrossRef]
- Mendoza-Cózatl, D.; Rangel-González, E.; Moreno-Sánchez, R. Simultaneous Cd2+, Zn2+, and Pb2+ uptake and accumulation by photosynthetic Euglena gracilis. Arch. Environ. Contam. Toxicol. 2006, 51, 521–528. [Google Scholar] [CrossRef]
- Clemens, S. Toxic metal accumulation, responses to exposure and mechanisms of tolerance in plants. Biochimie 2006, 88, 1707–1719. [Google Scholar] [CrossRef]
- Leal-Alvarado, D.A.; Espadas, F.; Sáenz-Carbonell, L.; Talavera, C. Lead accumulation reduces photosynthesis in the lead hyper-accumulator Salvinia minima Baker by affecting the cell membrane and inducing stomatal closure. Aquatic Toxicol. 2016, 171, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Gomes, S.M.D.S.; De Lima, V.L.A.; De Souza, A.P.; Do Nascimento, J.J.V.R.; Do Nascimento, E.S. Chloroplast pigments as indicators of lead stress. Eng. Agríc. 2014, 34, 877–884. [Google Scholar] [CrossRef] [Green Version]
- Ramel, F.; Mialoundama, A.S.; Havaux, M. Nomenzymic carotenoid oxidation and photooxidative stress signalling in plants. J. Exp. Bot. 2013, 64, 799–805. [Google Scholar] [CrossRef] [PubMed]
- Munné-Bosch, S.; Queval, G.; Foyer, C.H. The impact of global change factors on redox signaling underpinning stress tolerance. Plant Physiol. 2013, 161, 5–19. [Google Scholar] [CrossRef] [Green Version]
- Dhir, B.; Sharmila, P.; Pardha-Saradhi, P. Photosynthetic performance of Salvinia natans exposed to chromium and zinc rich wastewater. Braz. J. Plant Physiol. 2008, 20, 61–70. [Google Scholar] [CrossRef]
- Chaney, R.L. Zinc Phytotoxicity. In Zinc in Soils and Plants; Robson, A.D., Ed.; Springer: Dordrecht, The Netherlands, 1993; Volume 55, pp. 135–150. [Google Scholar] [CrossRef]
- Broadley, M.R.; White, P.J.; Hammond, J.P.; Zelko, I.; Lux, A. Zinc in plants. New Phytol. 2007, 173, 677–702. [Google Scholar] [CrossRef]
- Prado, C.; Ponce, S.C.; Pagano, E.; Prado, F.E.; Rosa, M. Differential physiological responses of two Salvinia species to hexavalent chromium at a glance. Aquatic Toxicol. 2016, 175, 213–221. [Google Scholar] [CrossRef]
- Michalak, A. Phenolic compounds and their antioxidant activity in plants growing under heavy metal stress. Polish J. Environ. Stud. 2006, 15, 523–530. [Google Scholar]
- Dhir, B.; Sharmila, P.; Pardha-Saradhi, P.; Sharma, P.; Kumar, R.; Mehta, D. Heavy metal induced physiological alterations in Salvinia natans. Ecotoxicol. Environ. Saf. 2011, 74, 1678–1684. [Google Scholar] [CrossRef]
- Hall, J.L. Cellular mechanisms for heavy metal detoxification and tolerance. J. Exp. Bot. 2002, 53, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Parihar, P.; Singh, R.; Singh, V.P.; Prasad, S.M. Heavy metal tolerance in plants: Role of transcriptomics, proteomics, metabolomics, and ionomics. Front. Plant Sci. 2016, 6, 1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Exposure Time (h) | 50 ± 2 µM | 100 ± 1 µM | ||||||
|---|---|---|---|---|---|---|---|---|
| Cd | Cu | Pb | Zn | Cd | Cu | Pb | Zn | |
| 0 | 0A | 0A | 0A | 0A | 0A | 0A | 0A | 0A |
| 2 | 27 ± 2bB | 46 ± 9cB | 50 ± 7cB | 29 ± 1bB | 19 ± 3aB | 42 ± 3cB | 66 ± 1dB | 15 ± 2aB |
| 4 | 52 ± 2bC | 62 ± 6cC | 82 ± 1dC | 49 ± 1bC | 25 ± 6aBC | 66 ± 7cBC | 81 ± 3dC | 33 ± 4aC |
| 6 | 56 ± 6bC | 76 ± 7cCD | 90 ± 2dD | 51 ± 1bC | 30 ± 7aC | 77 ± 9cCD | 86 ± 2cdD | 42 ± 7abC |
| 10 | 69 ± 3cD | 90 ± 8deDE | 97 ± 1eE | 67 ± 7cD | 37 ± 3aC | 84 ± 6dD | 94 ± 1eE | 59 ± 1bD |
| 24 | 72 ± 1bDE | 99 ± 2cE | 98 ± 1cE | 72 ± 3bDE | 50 ± 4aD | 98 ± 1cE | 96 ± 1cE | 69 ± 3bDE |
| 48 | 79 ± 4cE | 100 ± 2dE | 99 ± 2dE | 77 ± 5cE | 56 ± 2aD | 99 ± 2dE | 97 ± 2dE | 70 ± 4bE |
| Metal (µM) | First-order Kinetics | Second-order Kinetics | |||
|---|---|---|---|---|---|
| K1 (1/h) | R2 | K2 (L/mg.h) | R2 | ||
| Cd | 50 ± 2 | 0.1163 | 0.9227 | 0.0577 | 0.9922 |
| 100 ± 1 | 0.0424 | 0.8921 | 0.0080 | 0.9932 | |
| Cu | 50 ± 2 | 0.1859 | 0.9936 | 1.3434 | 0.8932 |
| 100 ± 1 | 0.1770 | 0.9905 | 0.6033 | 0.8842 | |
| Pb | 50 ± 2 | 0.3986 | 0.9907 | 0.1991 | 0.9340 |
| 100 ± 1 | 0.3173 | 0.9950 | 0.0945 | 0.8399 | |
| Zn | 50 ± 2 | 0.1020 | 0.9413 | 0.0939 | 0.9985 |
| 100 ± 1 | 0.0928 | 0.9362 | 0.0230 | 0.9946 | |
| Physiological Parameters | Control | Cd (µM) | Cu (µM) | Pb (µM) | Zn (µM) | ||||
|---|---|---|---|---|---|---|---|---|---|
| 50 ± 2 | 100 ± 1 | 50 ± 2 | 100 ± 1 | 50 ± 2 | 100 ± 1 | 50 ± 2 | 100 ± 1 | ||
| chl a (µg/g FW) | 156 ± 7a (100%) | 151 ± 15a (97%) | 160 ± 19a (102%) | 138 ± 24ab (88%) | 102 ± 16b (65%) | 123 ± 10b (79%) | 113 ± 5b (72%) | 156 ± 18a (100%) | 166 ± 36a (106%) |
| chl b (µg/g FW) | 109 ± 6a (100%) | 92 ± 8a (84%) | 92 ±7a (84%) | 78 ± 12b (71%) | 65 ± 9b (62%) | 82 ± 7b (75%) | 81 ± 9b (74%) | 109 ± 14a (100%) | 105 ± 22a (101%) |
| chl total (µg/g FW) | 261 ± 14a (100%) | 242 ± 24a (93%) | 257 ± 11a (98%) | 198 ± 35b (76%) | 167 ± 25b (64%) | 205 ± 16b (78%) | 194 ± 14b (74%) | 265 ± 25a (102%) | 271 ± 41a (104%) |
| Carotenoids (µg/g FW) | 35 ± 2a (100%) | 43 ± 3b (123%) | 43 ± 2b (123%) | 37 ± 2a (106%) | 28 ± 2b (80%) | 36 ± 3a (103%) | 37 ± 2a (106%) | 34 ± 2a (97%) | 38 ± 4a (108%) |
| Flavonoids (Abs/g FW) | 87 ± 5a (100%) | 89 ± 4a (102%) | 82 ±8a (94%) | 91 ± 8a (105%) | 84 ± 4a (97%) | 95 ± 8a (109%) | 86 ± 7a (99%) | 93 ± 7a (107%) | 83 ± 7a (95%) |
| Anthocyanins (Abs/g FW) | 560 ± 46a (100%) | 584 ± 45a (104%) | 549 ± 75a (98%) | 545 ± 8a (97%) | 596 ± 37a (106%) | 603 ± 78a (108%) | 566 ± 45a (101%) | 610 ± 60a (109%) | 581 ± 50a (104%) |
| Carbohydrates (mg/g FW) | 42 ± 1a (100%) | 43 ± 3a (102%) | 41 ± 5a (98%) | 40 ± 7a (95%) | 38 ± 9a (90%) | 46 ± 2a (109%) | 37 ± 2a (88%) | 41 ± 7a (98%) | 40 ± 7a (95%) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Emiliani, J.; Llatance Oyarce, W.G.; Bergara, C.D.; Salvatierra, L.M.; Novo, L.A.B.; Pérez, L.M. Variations in the Phytoremediation Efficiency of Metal-polluted Water with Salvinia biloba: Prospects and Toxicological Impacts. Water 2020, 12, 1737. https://doi.org/10.3390/w12061737
Emiliani J, Llatance Oyarce WG, Bergara CD, Salvatierra LM, Novo LAB, Pérez LM. Variations in the Phytoremediation Efficiency of Metal-polluted Water with Salvinia biloba: Prospects and Toxicological Impacts. Water. 2020; 12(6):1737. https://doi.org/10.3390/w12061737
Chicago/Turabian StyleEmiliani, Julia, Wendi G. Llatance Oyarce, C. Daniela Bergara, Lucas M. Salvatierra, Luís A. B. Novo, and Leonardo M. Pérez. 2020. "Variations in the Phytoremediation Efficiency of Metal-polluted Water with Salvinia biloba: Prospects and Toxicological Impacts" Water 12, no. 6: 1737. https://doi.org/10.3390/w12061737