Taxonomic and Functional Responses of Benthic Macroinvertebrate Communities to Hydrological and Water Quality Variations in a Heavily Regulated River


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
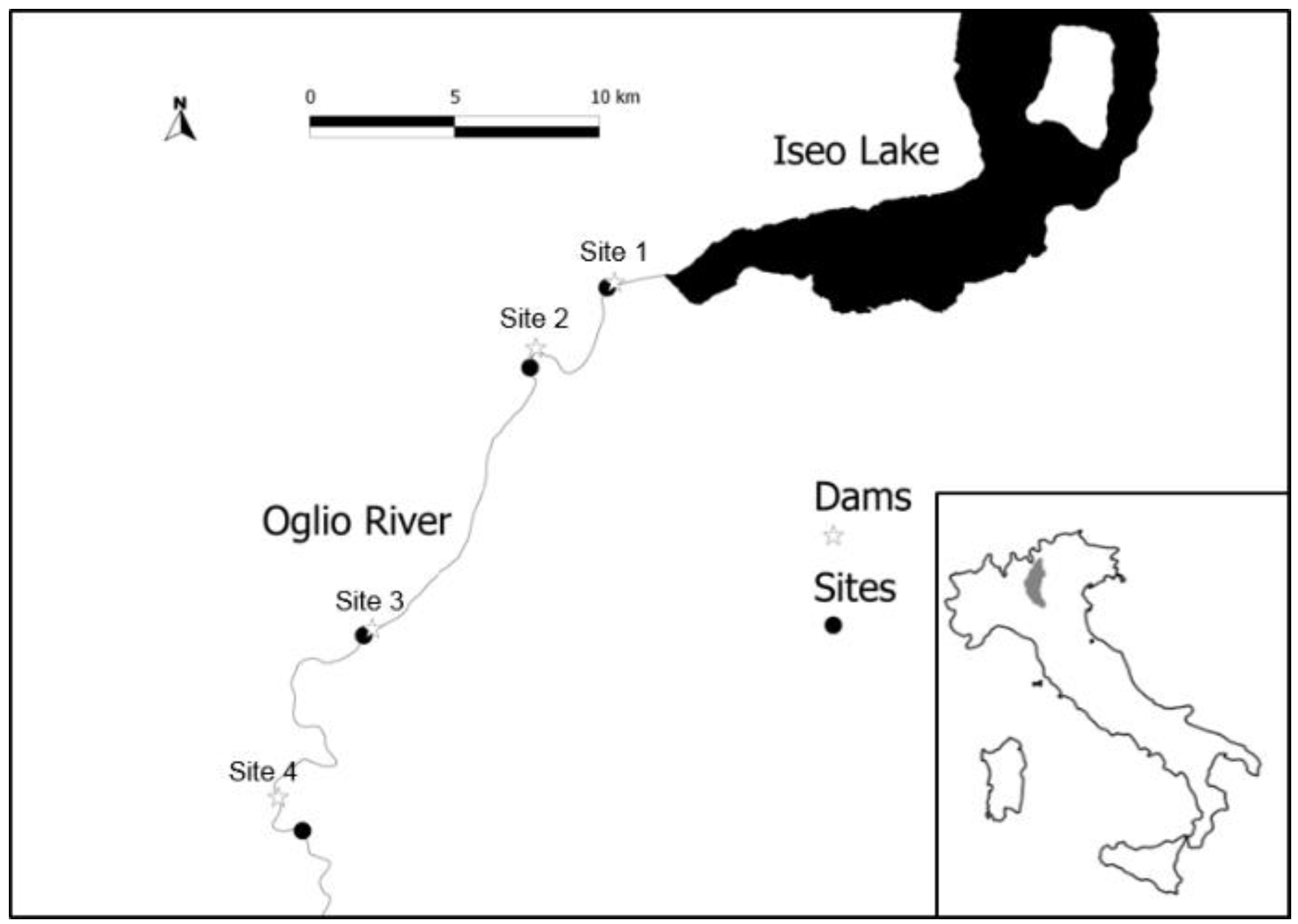
2.1. Study Area
2.2. Sampling Activity: Biological, Hydrological and Water Quality Data
2.3. Macroinvertebrate Metrics and Indices: Taxonomic and Functional Approaches
2.4. Statistical Analysis
3. Results
3.1. Hydrology and Water Quality Data
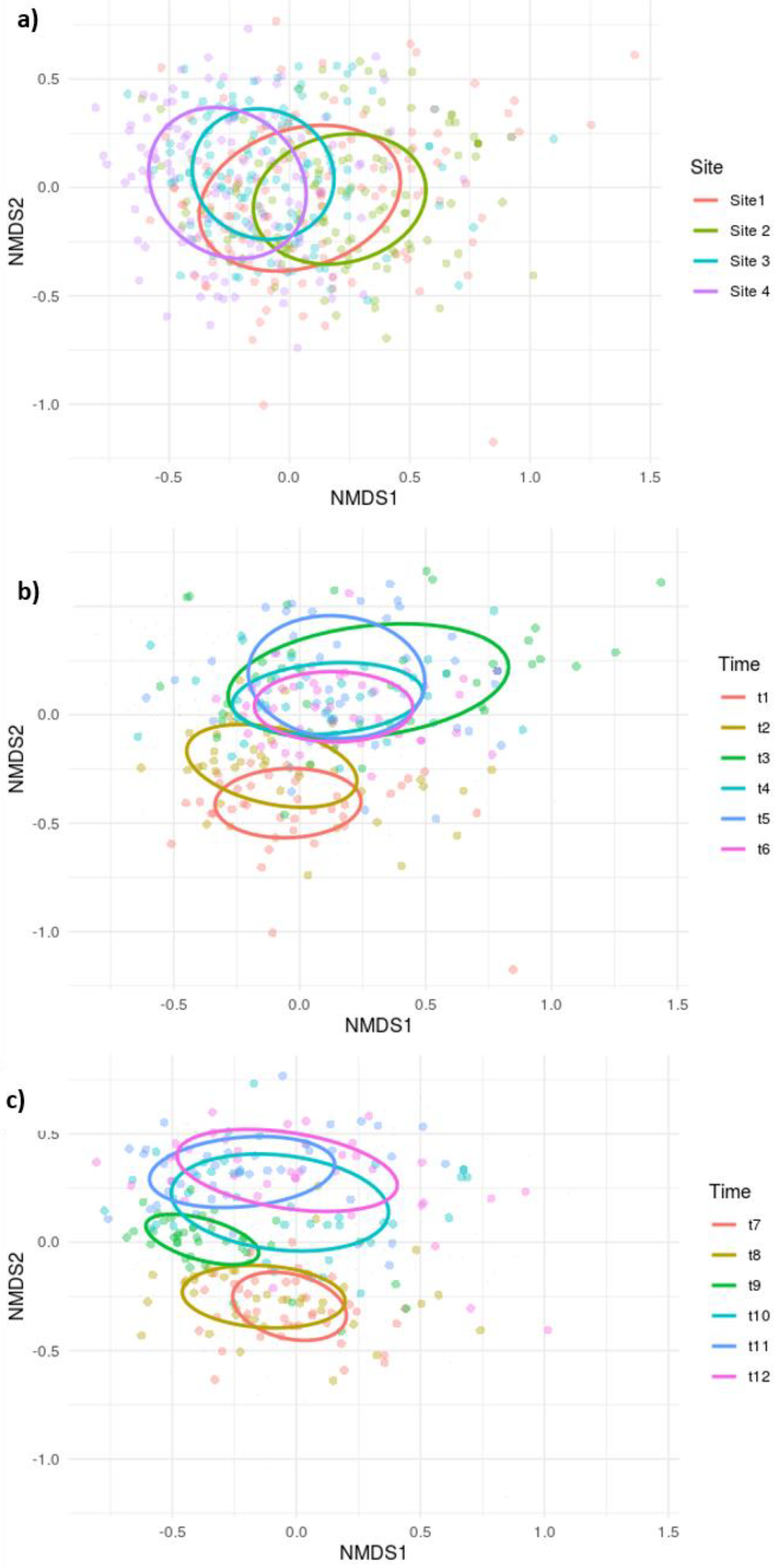
3.2. Macroinvertebrate Community: Responses, Ordinations and Indicator Taxa
4. Discussion
4.1. Macroinvertebrate Responses: The Importance of Fixed and Random Effects
4.2. Differences in the Similarity: Cross-Site Comparison
4.3. Remarks and Implications for River Monitoring and Management
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nilsson, C.; Reidy, C.A.; Dynesius, M.; Revenga, C. Fragmentation and Flow Regulation of the World’s Large River Systems. Science 2005, 308, 405–408. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, I.P.; Diamond, M.; Gurnell, A.M.; Hall, K.A.; Jenkins, A.; Milner, N.J.; Naylor, L.A.; Sear, D.A.; Woodward, G.; Ormerod, S.J. Integrating ecology with hydromorphology: A priority for river science and management. Aquat. Conserv. Mar. Freshw. Ecosyst. 2009, 19, 113–125. [Google Scholar] [CrossRef]
- Zarfl, C.; Lumsdon, A.E.; Berlekamp, J.; Tydecks, L.; Tockner, K. A global boom in hydropower dam construction. Aquat. Sci. 2015, 77, 161–170. [Google Scholar] [CrossRef]
- Richter, B.D.; Mathews, R.; Harrison, D.L.; Wigington, R. Ecologically Sustainable Water Management: Managing River Flows for Ecological Integrity. Ecol. Appl. 2003, 13, 206–224. [Google Scholar] [CrossRef]
- Arthington, A.H.; Naiman, R.J.; Mcclain, M.E.; Nilsson, C. Preserving the biodiversity and ecological services of rivers: New challenges and research opportunities. Freshw. Biol. 2010, 55, 1–16. [Google Scholar]
- Schlatter, K.J.; Grabau, M.R.; Shafroth, P.B.; Zamora-Arroyo, F. Integrating active restoration with environmental flows to improve native riparian tree establishment in the Colorado River Delta. Ecol. Eng. 2017, 106, 661–674. [Google Scholar] [CrossRef]
- Davies, P.M.; Naiman, R.J.; Warfe, D.M.; Pettit, N.E.; Arthington, A.H.; Bunn, S.E. Flow–ecology relationships: Closing the loop on effective environmental flows. Mar. Freshw. Res. 2014, 65, 133–141. [Google Scholar] [CrossRef]
- Bolpagni, R.; Racchetti, E.; Laini, A. Fragmentation and groundwater supply as major drivers of algal and plant diversity and relative cover dynamics along a highly modified lowland river. Sci. Total Environ. 2016, 568, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Bonada, N.; Prat, N.; Resh, V.H.; Statzner, B. Developments in aquatic insects biomonitoring: A Comparative Analysis of Recent Approaches. Annu. Rev. Entomol. 2006, 51, 495–523. [Google Scholar] [CrossRef]
- Friberg, N.; Skriver, J.; Larsen, S.E.; Pedersen, M.L.; Buffagni, A. Stream macroinvertebrate occurrence along gradients in organic pollution and eutrophication. Freshw. Biol. 2010, 55, 1405–1419. [Google Scholar] [CrossRef]
- Bruno, D.; Belmar, O.; Sánchez-Fernández, D.; Guareschi, S.; Millán, A.; Velasco, J. Responses of Mediterranean aquatic and riparian communities to human pressures at different spatial scales. Ecol. Indic. 2014, 45, 456–464. [Google Scholar] [CrossRef]
- Buss, D.F.; Carlisle, D.M.; Chon, T.-S.; Culp, J.; Harding, J.S.; Keizer-Vlek, H.E.; Robinson, W.A.; Strachan, S.; Thirion, C.; Hughes, R.M. Stream biomonitoring using macroinvertebrates around the globe: A comparison of large-scale programs. Environ. Monit. Assess. 2015, 187, 4132. [Google Scholar] [CrossRef] [PubMed]
- Salmaso, F.; Crosa, G.; Espa, P.; Gentili, G.; Quadroni, S.; Zaccara, S. Benthic macroinvertebrates response to water management in a lowland river: Effects of hydro-power vs irrigation off-stream diversions. Environ. Monit. Assess. 2017, 190, 33. [Google Scholar] [CrossRef] [PubMed]
- Céréghino, R.; Lavandier, P. Influence of hydropeaking on the distribution and larval development of the Plecoptera from a mountain stream. Regul. Rivers Res. Manag. 1998, 14, 297–309. [Google Scholar] [CrossRef]
- Stubbington, R.; Wood, P.J.; Boulton, A.J. Low flow controls on benthic and hyporheic macroinvertebrate assemblages during supra-seasonal drought. Hydrol. Process. 2009, 23, 2252–2263. [Google Scholar] [CrossRef] [Green Version]
- Dolédec, S.; Lamouroux, N.; Fuchs, U.; Mérigoux, S. Modelling the hydraulic preferences of benthic macroinvertebrates in small European streams. Freshw. Biol. 2007, 52, 145–164. [Google Scholar] [CrossRef]
- Brooks, A.J.; Haeusler, T.; Reinfelds, I.; Williams, S. Hydraulic microhabitats and the distribution of macroinvertebrate assemblages in riffles. Freshw. Biol. 2005, 50, 331–344. [Google Scholar] [CrossRef]
- Camargo, J.A.; Voelz, N.J. Biotic and Abiotic Changes along the Recovery Gradient of Two Impounded Rivers with Different Impoundment Use. Environ. Monit. Assess. 1998, 50, 143–158. [Google Scholar] [CrossRef]
- Olden, J.D.; Naiman, R.J. Incorporating thermal regimes into environmental flows assessments: Modifying dam operations to restore freshwater ecosystem integrity. Freshw. Biol. 2010, 55, 86–107. [Google Scholar] [CrossRef]
- Nilsson, C.; Renöfält, B. Linking Flow Regime and Water Quality in Rivers: A Challenge to Adaptive Catchment Management. Ecol. Soc. 2008, 13, 18. [Google Scholar] [CrossRef]
- Mouillot, D.; Graham, N.A.J.; Villéger, S.; Mason, N.W.H.; Bellwood, D.R. A functional approach reveals community responses to disturbances. Trends Ecol. Evol. 2013, 28, 167–177. [Google Scholar] [CrossRef]
- Bruno, D.; Gutiérrez-Cánovas, C.; Velasco, J.; Sánchez-Fernández, D. Functional redundancy as a tool for bioassessment: A test using riparian vegetation. Sci. Total Environ. 2016, 566–567, 1268–1276. [Google Scholar] [CrossRef] [PubMed]
- Ruhi, A.; Dong, X.; McDaniel, C.H.; Batzer, D.P.; Sabo, J.L. Detrimental effects of a novel flow regime on the functional trajectory of an aquatic invertebrate metacommunity. Glob. Chang. Biol. 2018, 24, 3749–3765. [Google Scholar] [CrossRef] [Green Version]
- Guareschi, S.; Laini, A.; Racchetti, E.; Bo, T.; Fenoglio, S.; Bartoli, M. How do hydromorphological constraints and regulated flows govern macroinvertebrate communities along an entire lowland river? Ecohydrology 2014, 7, 366–377. [Google Scholar] [CrossRef]
- Buffagni, A.; Erba, S. Parte A—Metodo di campionamento per i fiumi guadabili. In Notiziario dei Metodi Analitici; Istituto di Ricerca Sulle Acque del Consiglio Nazionale Delle Ricerche: Roma, Italy, 2007. [Google Scholar]
- Bartoli, M.; Racchetti, E.; Delconte, C.A.; Sacchi, E.; Soana, E.; Laini, A.; Longhi, D.; Viaroli, P. Nitrogen balance and fate in a heavily impacted watershed (Oglio River, Northern Italy): In quest of the missing sources and sinks. Biogeosciences 2012, 9, 361–373. [Google Scholar] [CrossRef]
- Lancaster, J.; Belyea, L.R. Defining the limits to local density: Alternative views of abundance–environment relationships. Freshw. Biol. 2006, 51, 783–796. [Google Scholar] [CrossRef]
- Tachet, H. Invertébrés d’eau Douce: Systématique, Biologie, Écologie; édition Revue et Augmentée Edizione; CNRS EDITIONS: Paris, France, 2010; ISBN 978-2-271-06945-0. [Google Scholar]
- Guareschi, S.; Laini, A.; Sánchez-Montoya, M.M. How do low-abundance taxa affect river biomonitoring? Exploring the response of different macroinvertebrate-based indices. J. Limnol. 2017, 76, 9–20. [Google Scholar] [CrossRef]
- Heino, J.; Soininen, J. Are higher taxa adequate surrogates for species-level assemblage patterns and species richness in stream organisms? Biol. Conserv. 2007, 137, 78–89. [Google Scholar] [CrossRef]
- Belmar, O.; Velasco, J.; Gutiérrez-Cánovas, C.; Mellado-Díaz, A.; Millán, A.; Wood, P.J. The influence of natural flow regimes on macroinvertebrate assemblages in a semiarid Mediterranean basin. Ecohydrology 2013, 6, 363–379. [Google Scholar] [CrossRef]
- American Public Health Association; American Water Works Association; Water Environment Federation. Standard Methods for the Examination of Water and Wastewater, 23th ed.; Water Environment Federation: Washington, DC, USA, 2017; ISBN 978-0-87553-287-5. [Google Scholar]
- APAT-CNR-IRSA. Manuale Linee Guida-Metodi Analitici per le Acque; APAT: Roma, Italy, 2003; ISBN 88-448-0083-7. [Google Scholar]
- Birk, S.; Bonne, W.; Borja, A.; Brucet, S.; Courrat, A.; Poikane, S.; Solimini, A.; van de Bund, W.; Zampoukas, N.; Hering, D. Three hundred ways to assess Europe’s surface waters: An almost complete overview of biological methods to implement the Water Framework Directive. Ecol. Indic. 2012, 18, 31–41. [Google Scholar] [CrossRef]
- Mondy, C.P.; Villeneuve, B.; Archaimbault, V.; Usseglio-Polatera, P. A new macroinvertebrate-based multimetric index (I2M2) to evaluate ecological quality of French wadeable streams fulfilling the WFD demands: A taxonomical and trait approach. Ecol. Indic. 2012, 18, 452–467. [Google Scholar] [CrossRef]
- Laini, A.; Bolpagni, R.; Cancellario, T.; Guareschi, S.; Racchetti, E.; Viaroli, P. Testing the response of macroinvertebrate communities and biomonitoring indices under multiple stressors in a lowland regulated river. Ecol. Indic. 2018, 90, 47–53. [Google Scholar] [CrossRef]
- Hawkes, H.A. Origin and development of the biological monitoring working party score system. Water Res. 1998, 32, 964–968. [Google Scholar] [CrossRef]
- Ministry of Agriculture, Fisheries and Food (MAGRAMA). Criterios de Seguimiento Y Evaluación del Estado de Las Aguas Superficiales Y Las Normas de Calidad Ambiental. Boletín Oficial del Estado-BOE, 219: 80582–80677; Ministerio de Agricultura, Alimentación y Medio Ambiente: Madrid, Spain, 2015. [Google Scholar]
- The UK Technical Advisory Group (UKTAG). UKTAG river assessment method, benthic invertebrate fauna. In Invertebrates (General Degradation): Whalley, Hawkes, Paisley & Trigg (WHPT). Metric in River Invertebrate Classification Tool (RICT); WFD-UKTAG: Scotland, UK, 2014. [Google Scholar]
- Villéger, S.; Mason, N.W.H.; Mouillot, D. New Multidimensional Functional Diversity Indices for a Multifaceted Framework in Functional Ecology. Ecology 2008, 89, 2290–2301. [Google Scholar] [CrossRef] [PubMed]
- Pillar, V.D.; Blanco, C.C.; Müller, S.C.; Sosinski, E.E.; Joner, F.; Duarte, L.D.S. Functional redundancy and stability in plant communities. J. Veg. Sci. 2013, 24, 963–974. [Google Scholar] [CrossRef]
- Hooper, D.U.; Chapin, F.S.; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of Biodiversity on Ecosystem Functioning: A Consensus of Current Knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Schmera, D.; Heino, J.; Podani, J.; Erős, T.; Dolédec, S. Functional diversity: A review of methodology and current knowledge in freshwater macroinvertebrate research. Hydrobiologia 2017, 787, 27–44. [Google Scholar] [CrossRef]
- Hevia, V.; Martín-López, B.; Palomo, S.; García-Llorente, M.; de Bello, F.; González, J.A. Trait-based approaches to analyze links between the drivers of change and ecosystem services: Synthesizing existing evidence and future challenges. Ecol. Evol. 2017, 7, 831–844. [Google Scholar] [CrossRef] [PubMed]
- Belmar, O.; Bruno, D.; Guareschi, S.; Mellado-Díaz, A.; Millán, A.; Velasco, J. Functional responses of aquatic macroinvertebrates to flow regulation are shaped by natural flow intermittence in Mediterranean streams. Freshw. Biol. 2019, 64, 1064–1077. [Google Scholar] [CrossRef]
- Zuur, A.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Statistics for Biology and Health; Springer: New York, NY, USA, 2009; ISBN 978-0-387-87457-9. [Google Scholar]
- Mccarthy, M.A. Bayesian Methods for Ecology, 1st ed.; Cambridge University Press: Cambridge, UK, 2007; ISBN 978-0-521-61559-4. [Google Scholar]
- Nakagawa, S.; Schielzeth, H. A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Dufrêne, M.; Legendre, P. Species Assemblages and Indicator Species: The Need for a Flexible Asymmetrical Approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- De Cáceres, M.D.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Naimi, B.; Hamm, N.A.S.; Groen, T.A.; Skidmore, A.K.; Toxopeus, A.G. Where is positional uncertainty a problem for species distribution modelling? Ecography 2014, 37, 191–203. [Google Scholar] [CrossRef]
- Hadfield, J.D. MCMC Methods for Multi-Response Generalized Linear Mixed Models: The MCMCglmm R Package. J. Stat. Softw. 2010, 33, 1–22. [Google Scholar] [CrossRef]
- Plummer, M.; Best, N.; Cowles, K.; Vines, K. CODA: Convergence diagnosis and output analysis for MCMC. R News 2006, 6, 7–11. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. Version. 2.5-5. CRAN, GitHub, US, 2019. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 12 May 2019).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Use R! Springer: New York, NY, USA, 2009; ISBN 978-0-387-98141-3. [Google Scholar]
- Fochetti, R.; Tierno De Figueroa, J.M. Plecoptera; Calderini: Milano, Italy, 2008; ISBN 978-88-506-5307-2. [Google Scholar]
- Poff, N.L.; Zimmerman, J.K.H. Ecological responses to altered flow regimes: A literature review to inform the science and management of environmental flows. Freshw. Biol. 2010, 55, 194–205. [Google Scholar] [CrossRef]
- Konrad, C.P.; Brasher, A.M.D.; May, J.T. Assessing streamflow characteristics as limiting factors on benthic invertebrate assemblages in streams across the western United States. Freshw. Biol. 2008, 53, 1983–1998. [Google Scholar] [CrossRef]
- Brittain, J.E.; Eikeland, T.J. Invertebrate drift—A review. Hydrobiologia 1988, 166, 77–93. [Google Scholar] [CrossRef]
- Fritz, K.M.; Dodds, W.K. Resistance and Resilience of Macroinvertebrate Assemblages to Drying and Flood in a Tallgrass Prairie Stream System. Hydrobiologia 2004, 527, 99–112. [Google Scholar] [CrossRef]
- Rice, S.P.; Buffin-Bélanger, T.; Lancaster, J.; Reid, I. 24 Movements of a macroinvertebrate (Potamophylax latipennis) across a gravel-bed substrate: Effects of local hydraulics and micro-topography under increasing discharge. Dev. Earth Surf. Process. 2007, 11, 637–659. [Google Scholar]
- Hart, D.D.; Clark, B.D.; Jasentuliyana, A. Fine-scale field measurement of benthic flow environments inhabited by stream invertebrates. Limnol. Oceanogr. 1996, 41, 297–308. [Google Scholar] [CrossRef]
- Lancaster, J.; Downes, B.J. Linking the hydraulic world of individual organisms to ecological processes: Putting ecology into ecohydraulics. River Res. Appl. 2010, 26, 385–403. [Google Scholar] [CrossRef]
- Bond, N.R.; Downes, B.J. The independent and interactive effects of fine sediment and flow on benthic invertebrate communities characteristic of small upland streams. Freshw. Biol. 2003, 48, 455–465. [Google Scholar] [CrossRef]
- Mesa, L.M. Effect of spates and land use on macroinvertebrate community in Neotropical Andean streams. Hydrobiologia 2010, 641, 85–95. [Google Scholar] [CrossRef]
- Suren, A.M.; Jowett, I.G. Effects of floods versus low flows on invertebrates in a New Zealand gravel-bed river. Freshw. Biol. 2006, 51, 2207–2227. [Google Scholar] [CrossRef]
- Gutiérrez-Cánovas, C.; Sánchez-Fernández, D.; Velasco, J.; Millán, A.; Bonada, N. Similarity in the difference: Changes in community functional features along natural and anthropogenic stress gradients. Ecology 2015, 96, 2458–2466. [Google Scholar] [CrossRef] [PubMed]
- Bruno, D.; Gutiérrez-Cánovas, C.; Sánchez-Fernández, D.; Velasco, J.; Nilsson, C. Impacts of environmental filters on functional redundancy in riparian vegetation. J. Appl. Ecol. 2016, 53, 846–855. [Google Scholar] [CrossRef]
- Bêche, L.A.; Mcelravy, E.P.; Resh, V.H. Long-term seasonal variation in the biological traits of benthic-macroinvertebrates in two Mediterranean-climate streams in California, USA. Freshw. Biol. 2006, 51, 56–75. [Google Scholar] [CrossRef]
- Munné, A.; Prat, N. Use of macroinvertebrate-based multimetric indices for water quality evaluation in Spanish Mediterranean rivers: An intercalibration approach with the IBMWP index. Hydrobiologia 2009, 628, 203. [Google Scholar] [CrossRef]
- Lecerf, A.; Usseglio-Polatera, P.; Charcosset, J.-Y.L.; Bracht, B.; Chauvet, E. Assessment of functional integrity of eutrophic streams using litter breakdown and benthic macroinvertebrates. Arch. Für Hydrobiol. 2006, 165, 105–126. [Google Scholar] [CrossRef]
- Berenzen, N.; Schulz, R.; Liess, M. Effects of chronic ammonium and nitrite contamination on the macroinvertebrate community in running water microcosms. Water Res. 2001, 35, 3478–3482. [Google Scholar] [CrossRef]
- Beketov, M. Different sensitivity of mayflies (Insecta, Ephemeroptera) to ammonia, nitrite and nitrate: Linkage between experimental and observational data. Hydrobiologia 2004, 528, 209–216. [Google Scholar] [CrossRef]
- Salmaso, N.; Morabito, G.; Garibaldi, L.; Mosello, R. Trophic Development of the Deep Lakes South of the Alps: A Comparative Analysis. Fundam. Appl. Limnol./Archiv Hydrobiol. 2007, 170, 177–196. [Google Scholar] [CrossRef]
- Beketov, M.A.; Liess, M. An indicator for effects of organic toxicants on lotic invertebrate communities: Independence of confounding environmental factors over an extensive river continuum. Environ. Pollut. 2008, 156, 980–987. [Google Scholar] [CrossRef] [PubMed]
- Canobbio, S.; Mezzanotte, V.; Sanfilippo, U.; Benvenuto, F. Effect of Multiple Stressors on Water Quality and Macroinvertebrate Assemblages in an Effluent-Dominated Stream. Water Air Soil Pollut. 2009, 198, 359–371. [Google Scholar] [CrossRef]
- Bolpagni, R.; Laini, A. Microhabitat patterns of soft-bodied benthic algae in a lowland river largely fed by groundwater. Fottea 2016, 16, 244–254. [Google Scholar] [CrossRef] [Green Version]
- Boulton, A.J.; Hancock, P.J. Rivers as groundwater-dependent ecosystems: A review of degrees of dependency, riverine processes and management implications. Aust. J. Bot. 2006, 54, 133–144. [Google Scholar] [CrossRef]
- Bonada, N.; Rieradevall, M.; Dallas, H.; Davis, J.; Day, J.; Figueroa, R.; Resh, V.H.; Prat, N. Multi-scale assessment of macroinvertebrate richness and composition in Mediterranean-climate rivers. Freshw. Biol. 2008, 53, 772–788. [Google Scholar] [CrossRef]
- Shmida, A.; Wilson, M.V. Biological Determinants of Species Diversity. J. Biogeogr. 1985, 12, 1–20. [Google Scholar] [CrossRef]
- Ćuk, R.; Miliša, M.; Atanacković, A.; Dekić, S.; Blažeković, L.; Žganec, K. Biocontamination of benthic macroinvertebrate assemblages in Croatian major rivers and effects on ecological quality assessment. Knowl. Manag. Aquat. Ecosyst. 2019, 420, 11. [Google Scholar] [CrossRef]
- Holt, C.R.; Pfitzer, D.; Scalley, C.; Caldwell, B.A.; Batzer, D.P. Macroinvertebrate Community Responses to Annual Flow Variation from River Regulation: An 11-Year Study. River Res. Appl. 2015, 31, 798–807. [Google Scholar] [CrossRef]
- Downes, B.J.; Hindell, J.S. What’s in a site? Variation in lotic macroinvertebrate density and diversity in a spatially replicated experiment. Austral Ecol. 2000, 25, 128–139. [Google Scholar] [CrossRef]
- Laini, A.; Vorti, A.; Bolpagni, R.; Viaroli, P. Small-scale variability of benthic macroinvertebrates distribution and its effects on biological monitoring. Ann. Limnol. Int. J. Limnol. 2014, 50, 211–216. [Google Scholar] [CrossRef] [Green Version]
- Burgazzi, G.; Guareschi, S.; Laini, A. The role of small-scale spatial location on macroinvertebrate community in an intermittent stream. Limnetica 2018, 37, 319–340. [Google Scholar]
- White, J.C.; Krajenbrink, H.J.; Hill, M.J.; Hannah, D.M.; House, A.; Wood, P.J. Habitat-specific invertebrate responses to hydrological variability, anthropogenic flow alterations, and hydraulic conditions. Freshw. Biol. 2019, 64, 555–576. [Google Scholar] [CrossRef]
- Arthington, A.H.; Kennen, J.G.; Stein, E.D.; Webb, J.A. Recent advances in environmental flows science and water management—Innovation in the Anthropocene. Freshw. Biol. 2018, 63, 1022–1034. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| SITE | 2014 | 2015 | ||
|---|---|---|---|---|
| Qmean | Range | Qmean | Range | |
| 1 | 30.1 | (3.0–163.5) | 6.4 | (3–9.3) |
| 2 | 50.3 | (17–161.3) | 19.1 | (6.2–58.1) |
| 3 | 41 | (7.1–166.8) | 10 | (4–27.5) |
| 4 | 51.6 | (10.2–178.4) | 12.5 | (6.9–34.5) |
| PARAMETER | 2014 | 2015 |
|---|---|---|
| O2 (%) | 105 ± 5.43 | 105 ± 8.10 |
| NH4+ (mg N L−1) | 0.06 ± 0.04 | 0.06 ± 0.04 |
| NO3− (mg N L−1) | 0.65 ± 0.27 | 0.54 ± 0.27 |
| TP (µg P L−1) | 16.3 ± 9.84 | 34.8 ± 21.0 |
| COD (mg O2 L−1) | 3.81 ± 5.31 | 4.00 ± 3.45 |
| T (°C) | 20.5 ± 2.10 | 21.6 ± 4.02 |
| pH | 8.20 ± 0.34 | 7.87 ± 0.38 |
| Electrical conductivity (µS cm−1) | 222 ± 24.0 | 230 ± 24.3 |
| VARIABLE | R2m | R2c | Site | Time | Site:Time |
|---|---|---|---|---|---|
| Family Richness | 0.15 | 0.40 | 0.02 | 0.10 | 0.13 |
| Abundance | 0.29 | 0.53 | 0.01 | 0.05 | 0.19 |
| EPT Richness | 0.18 | 0.60 | 0.24 | 0.04 | 0.14 |
| ASPT | 0.11 | 0.67 | 0.50 | 0.00 | 0.06 |
| Functional richness | 0.09 | 0.23 | 0.00 | 0.11 | 0.02 |
| Functional redundancy | 0.13 | 0.54 | 0.17 | 0.04 | 0.21 |
| COVARIATE | SITE1 | SITE2 | SITE3 | SITE4 |
|---|---|---|---|---|
| Qcv | 0.17 *** | 0.03 | 0.05 | 0.06 * |
| Qmin | 0.25 *** | 0.09 ** | 0.07 ** | 0.14 *** |
| % O2 | 0.17 *** | 0.08 ** | 0.19 *** | 0.19 *** |
| NH4+ | 0.12 ** | 0.36 *** | 0.32 *** | 0.07 * |
| NO3− | 0.20 *** | 0.50 *** | 0.73 *** | 0.22 *** |
| TP | 0.03 | 0.02 | 0.04 | 0.05 * |
| Temperature | 0.29 *** | 0.45 *** | 0.52 *** | 0.39 *** |
| pH | 0.12 *** | 0.02 | 0.01 | 0.02 |
| Site 1 | ||||
| Order | Family | stat | p Value | |
| Veneroida | Dreissenidae | 0.870 | 0.0001 | *** |
| Basommatophora | Planorbidae | 0.767 | 0.0036 | ** |
| Diptera | Simuliidae | 0.744 | 0.0277 | * |
| Basommatophora | Acroloxidae | 0.738 | 0.0002 | *** |
| Anthoathecata | Hydridae | 0.737 | 0.0446 | * |
| Amphipoda | Gammaridae | 0.682 | 0.0115 | * |
| Diptera | Psychodidae | 0.624 | 0.0154 | * |
| Site 2 | ||||
| Order | Family | stat | p Value | |
| Trichoptera | Psychomyiidae | 0.646 | 0.0396 | * |
| Site 3 | ||||
| Order | Family | stat | p Value | |
| Trombidiformes | Hydracarina | 0.697 | 0.0001 | *** |
| Diptera | Limoniidae | 0.694 | 0.0206 | * |
| Isopoda | Asellidae | 0.661 | 0.0206 | * |
| Diptera | Chironomidae | 0.645 | 0.0226 | * |
| Trichoptera | Hydropsychidae | 0.622 | 0.048 | * |
| Site 4 | ||||
| Order | Family | stat | p Value | |
| Plecoptera | Leuctridae | 0.871 | 0.0001 | *** |
| Ephemeroptera | Ephemerellidae | 0.763 | 0.0057 | ** |
| Ephemeroptera | Heptageniidae | 0.746 | 0.0025 | ** |
| Veneroida | Corbiculidae | 0.715 | 0.0013 | ** |
| Arhynchobdellida | Erpobdellidae | 0.684 | 0.0036 | ** |
| Littorinimorpha | Hydrobiidae | 0.671 | 0.0052 | ** |
| Ephemeroptera | Caenidae | 0.665 | 0.0055 | ** |
| Hemiptera | Aphelocheiridae | 0.662 | 0.0063 | ** |
| Coleoptera | Elmidae | 0.649 | 0.0178 | * |
| Veneroida | Sphaeriidae | 0.648 | 0.0241 | * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laini, A.; Viaroli, P.; Bolpagni, R.; Cancellario, T.; Racchetti, E.; Guareschi, S. Taxonomic and Functional Responses of Benthic Macroinvertebrate Communities to Hydrological and Water Quality Variations in a Heavily Regulated River. Water 2019, 11, 1478. https://doi.org/10.3390/w11071478
Laini A, Viaroli P, Bolpagni R, Cancellario T, Racchetti E, Guareschi S. Taxonomic and Functional Responses of Benthic Macroinvertebrate Communities to Hydrological and Water Quality Variations in a Heavily Regulated River. Water. 2019; 11(7):1478. https://doi.org/10.3390/w11071478
Chicago/Turabian StyleLaini, Alex, Pierluigi Viaroli, Rossano Bolpagni, Tommaso Cancellario, Erica Racchetti, and Simone Guareschi. 2019. "Taxonomic and Functional Responses of Benthic Macroinvertebrate Communities to Hydrological and Water Quality Variations in a Heavily Regulated River" Water 11, no. 7: 1478. https://doi.org/10.3390/w11071478