Diversity of Bacteria in Lakes with Different Chlorophyll Content and Investigation of Their Respiratory Activity through a Long-Term Microcosm Experiment



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling—In Situ Abiotic and Biotic Parameters
2.2. Bacterial Diversity
2.3. Microcosm Experiments
3. Results
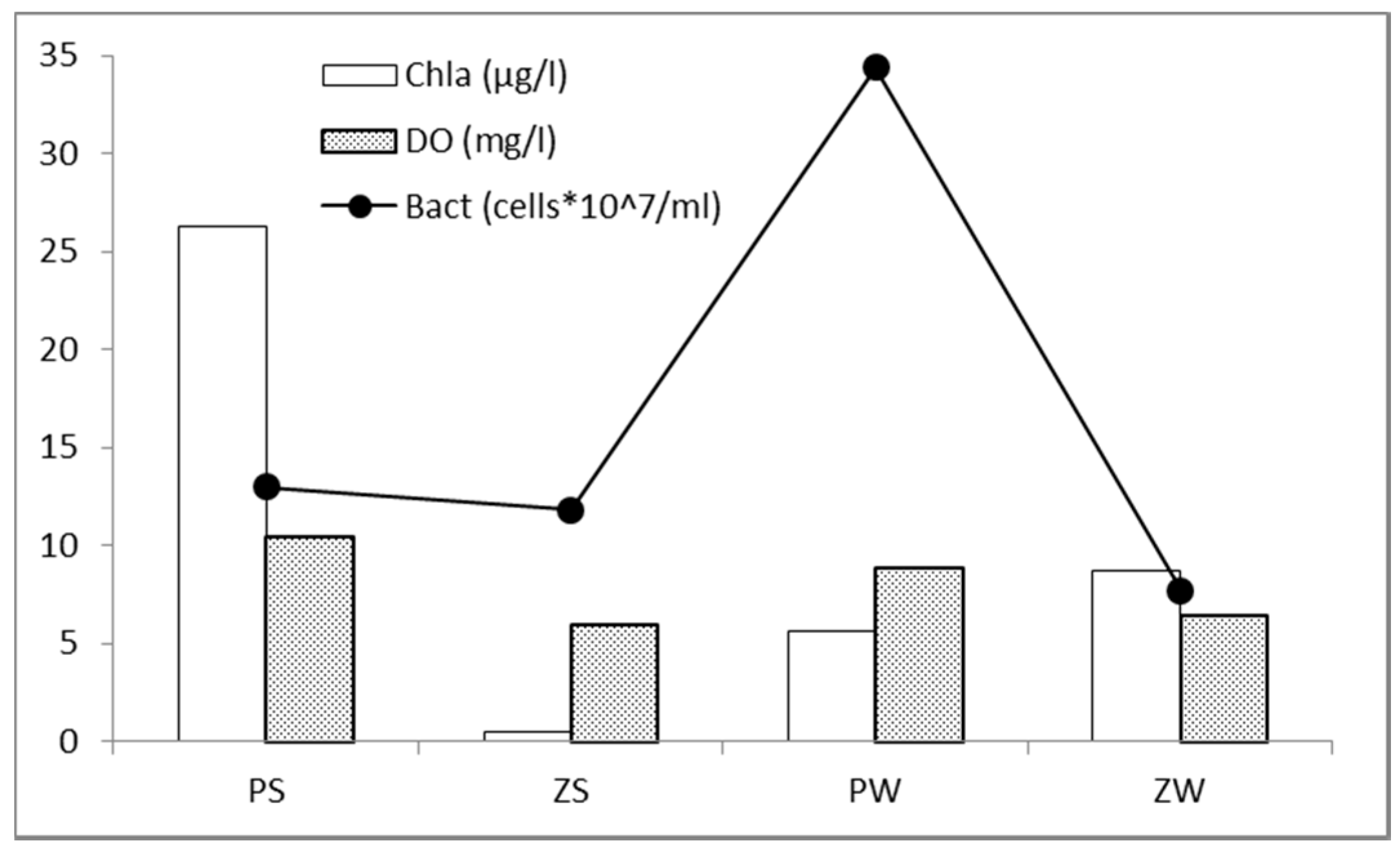
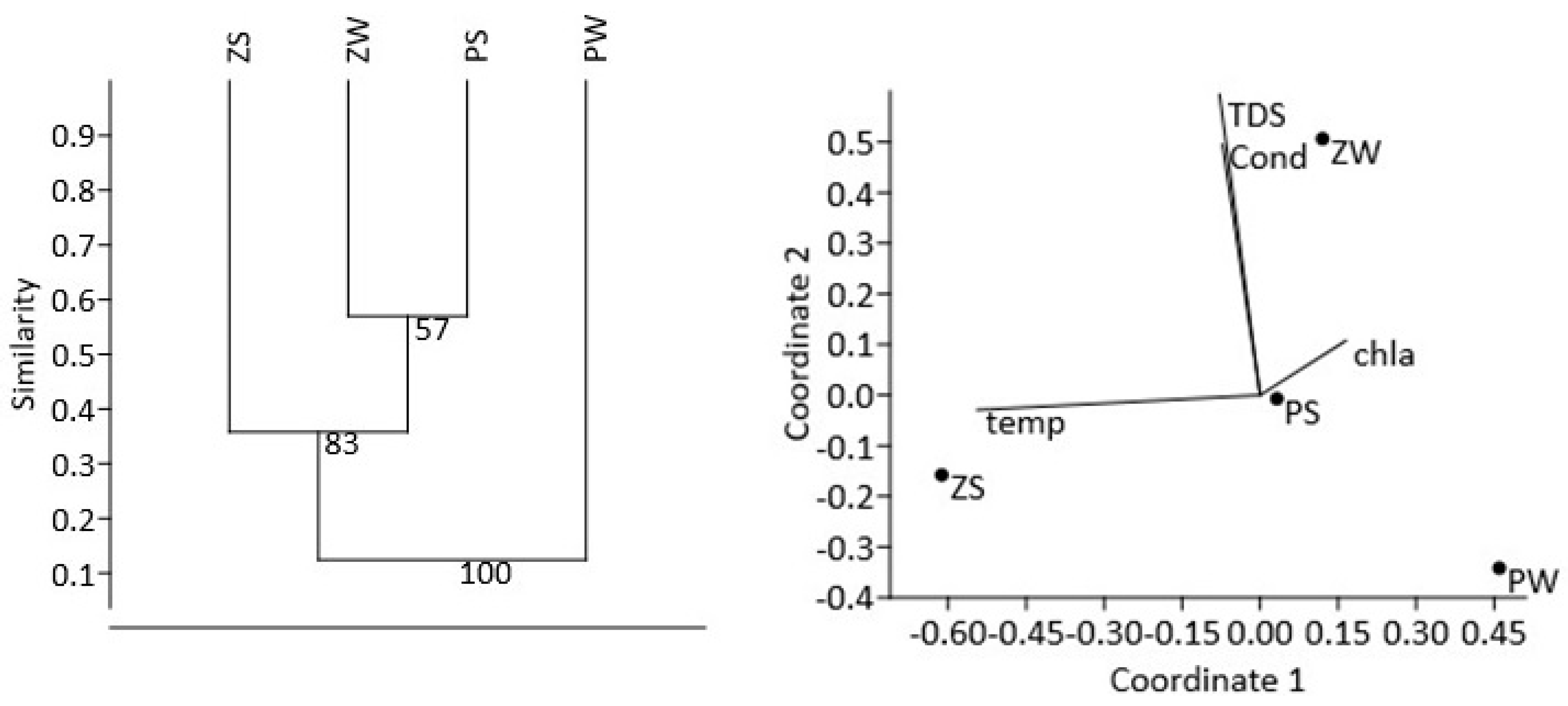
3.1. In Situ Abiotic and Biotic Parameters
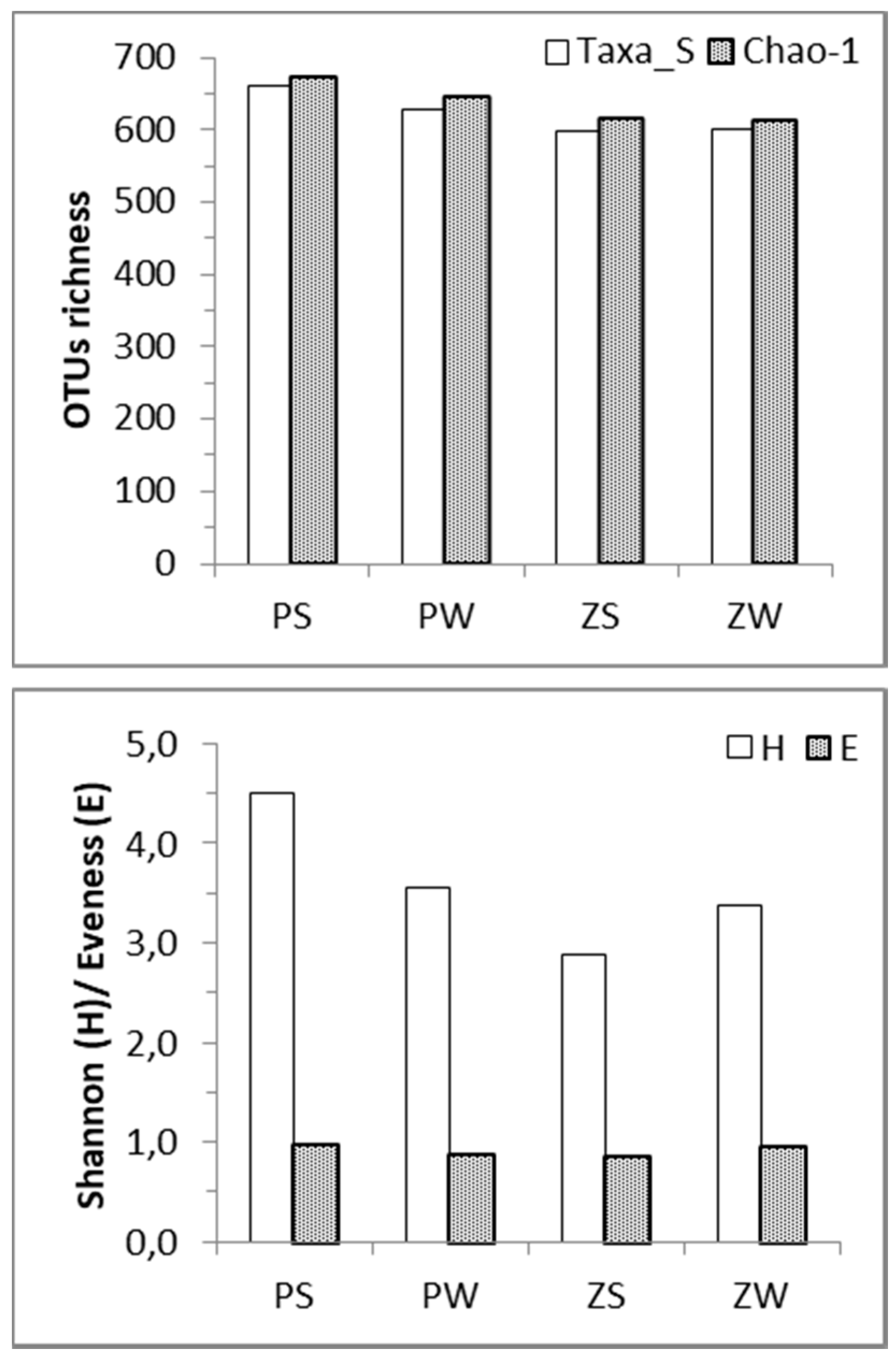
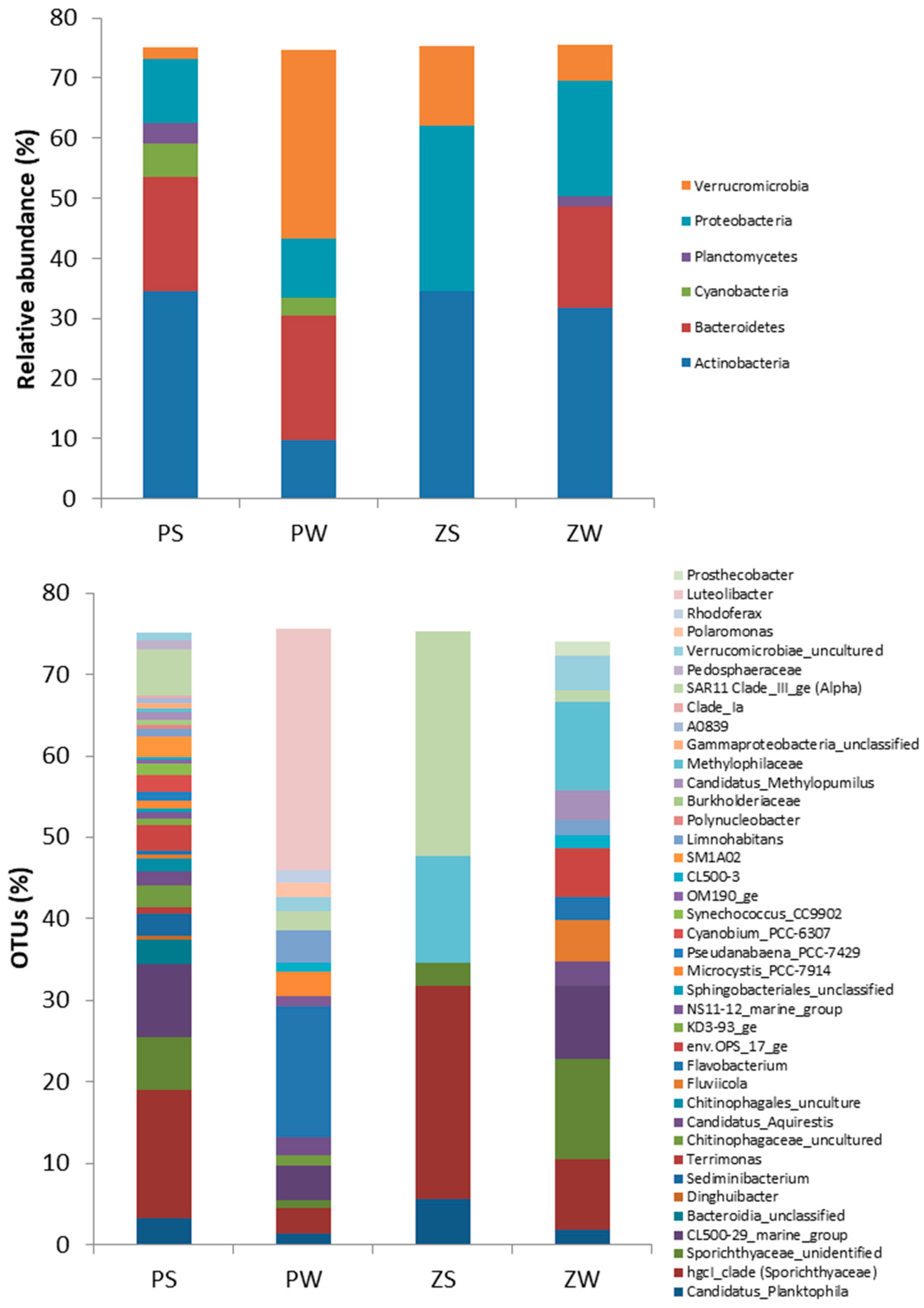
3.2. Bacterial Diversity
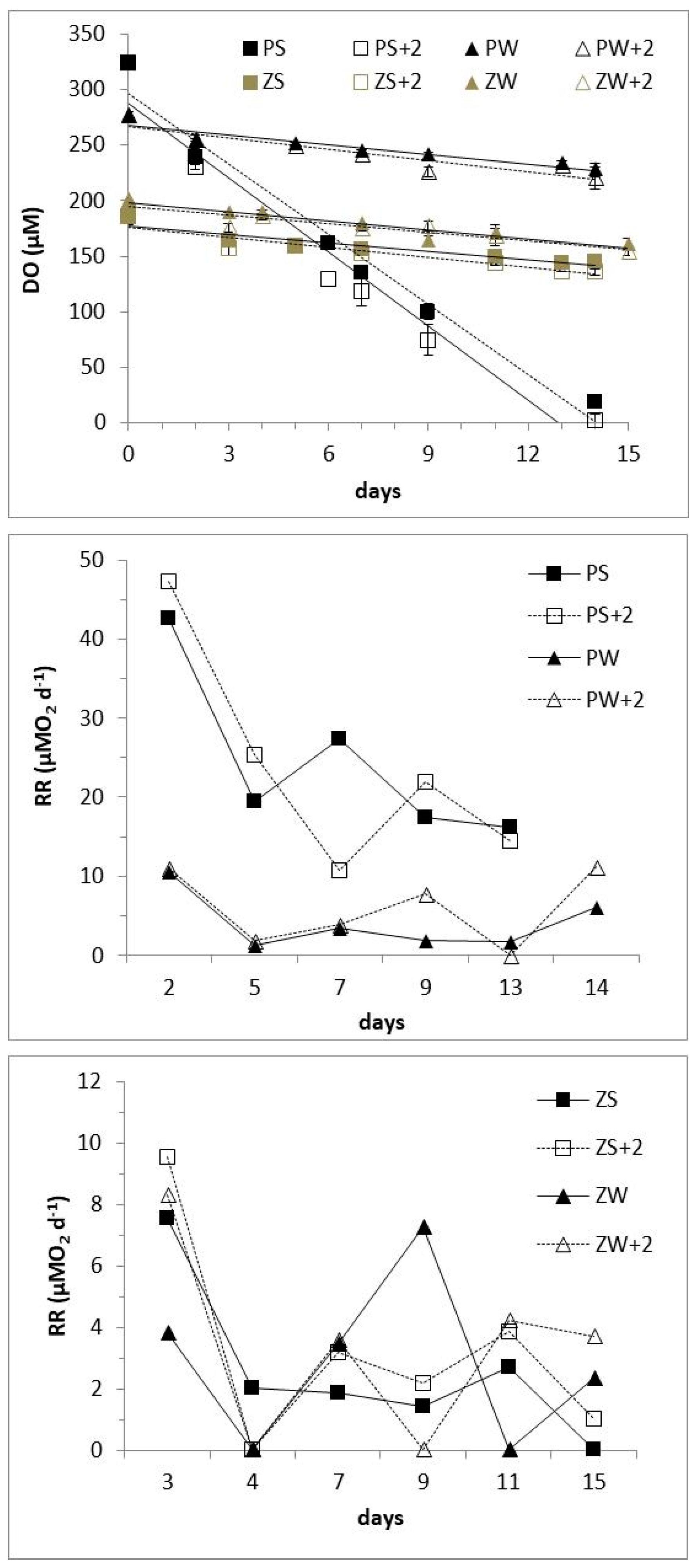
3.3. Microcosm Experiments
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Azam, F.; Fenchel, T.; Field, J.G.; Graf, J.S.; Meyer-Rei, L.-A.; Thingstad, F. The Ecological Role of Water-Column Microbes in the Sea. Mar. Ecol. Prog. Ser. 1983, 10, 257–263. [Google Scholar] [CrossRef]
- Sakamoto, C.; Yamaguchi, N.; Nasu, M. Rapid and simple quantification of bacterial cells by using a microfluidic device. Appl. Environ. Microbiol. 2005, 71, 1117–1121. [Google Scholar] [CrossRef] [PubMed]
- Comte, J.; Fauteux, L.; del Giorgio, P.A. Links between metabolic plasticity and functional redundancy in freshwater bacterioplankton communities. Front. Microbiol. 2013, 4, 112. [Google Scholar] [CrossRef] [PubMed]
- Pomeroy, L.R.; Sheldon, J.E.; Sheldon, W.M. Changes in Bacterial Numbers and Leucine Assimilation during Estimations of Microbial Respiratory Rates in Seawater by the Precision Winkler Method. Appl. Environ. Microbiol. 1994, 60, 328–332. [Google Scholar] [PubMed]
- Smith, E.M.; Prairie, Y.T. Bacterial metabolism and growth efficiency in lakes: The importance of phosphorus availability. Limnol. Oceanogr. 2004, 49, 137–147. [Google Scholar] [CrossRef] [Green Version]
- French, T.D.; Petticrew, E.L. Chlorophyll a seasonality in four shallow eutrophic lakes (northern British Columbia, Canada) and the critical roles of internal phosphorus loading and temperature. Hydrobiologia 2007, 575, 285–299. [Google Scholar] [CrossRef]
- OECD. Eutrophication of Waters. Monitoring, Assessment and Control; Organisation for Economic Co-Operation and Development: Paris, France, 1982; p. 154. [Google Scholar]
- Tsola, S.L.; Renta, C.; Macingo, S.C.; Karayanni, H. Investigating size-fractionated chlorophyll a and phycocyanin temporal variations in a highly eutrophic lake and its isolated karstic springs. Oceanol. Hydrobiol. Stud. 2018, 47, 118–127. [Google Scholar] [CrossRef]
- Bird, D.F.; Kalff, J. Empirical relationship between bacterial abundance and chlorophyll concentration in fresh and marine waters. Can. J. Fish. Aquat. Sci. 1984, 41, 1015–1023. [Google Scholar] [CrossRef]
- Inyang, A.A.E.; Dan, M. Bacterial Abundance—Chlorophyll a Concentration Relationships in Cross River Basin, Southeastern Nigeria: An Evaluation of Empirical Bacterial Abundance—Chlorophyll a Models Using a Multivariate Analysis. J. Water Resour. Ocean Sci. 2015, 4, 72–85. [Google Scholar]
- Muylaert, K.; Van Der Gucht, K.; Vloemans, N.; Meester, L.D.; Gillis, M.; Vyverman, W. Relationship between bacterial community composition and bottom-up versus top-down variables in four eutrophic shallow lakes. Appl. Environ. Microbiol. 2002, 68, 4740–4750. [Google Scholar] [CrossRef] [PubMed]
- Akins, L.; Ayayee, P.; Leff, L.G. Composition and diversity of cyanobacteria-associated and free-living bacterial communities during cyanobacterial blooms. Ann. Microbiol. 2018, 68, 493–503. [Google Scholar] [CrossRef]
- Louati, I.; Pascault, N.; Debroas, D.; Bernard, C.; Humbert, J.F.; Leloup, J. Structural Diversity of Bacterial Communities Associated with Bloom-Forming Freshwater Cyanobacteria Differs According to the Cyanobacterial Genus. PLoS ONE 2015, 10, e0140614. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Shen, H.; Chen, J.; Xie, P.; Yang, X.; Tao, M.; Ma, Z.; Qi, M. Phytoplankton community succession shaping bacterioplankton community composition in Lake Taihu, China. Water Res. 2011, 45, 4169–4182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ylla, I.; Peter, H.; Romani, A.M.; Tranvik, L.J. Different diversity-functioning relationship in lake and stream bacterial communities. FEMS Microbiol. Ecol. 2013, 85, 95–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keshri, J.; Pradeep Ram, A.S.; Nana, P.A.; Sime-Ngando, T. Taxonomical Resolution and Distribution of Bacterioplankton along the Vertical Gradient Reveals Pronounced Spatiotemporal Patterns in Contrasted Temperate Freshwater Lakes. Microb. Ecol. 2018, 76, 372–386. [Google Scholar] [CrossRef] [PubMed]
- De Figueiredo, D.R.; Pereira, M.J.; Moura, A.; Silva, L.; Barrios, S.; Fonseca, F.; Henriques, I.; Correia, A. Bacterial community composition over a dry winter in meso- and eutrophic Portuguese water bodies. FEMS Microbiol. Ecol. 2007, 59, 638–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llíros, M.; Inceoglu, O.; Garcia-Armisen, T.; Anzil, A.; Leporcq, B.; Pigneur, L.M.; Viroux, L.; Darchambeau, F.; Descy, J.P.; Servais, P. Bacterial community composition in three freshwater reservoirs of different alkalinity and trophic status. PLoS ONE 2014, 9, e116145. [Google Scholar] [CrossRef] [PubMed]
- Newton, R.J.; McLellan, S.L. A unique assemblage of cosmopolitan freshwater bacteria and higher community diversity differentiate an urbanized estuary from oligotrophic Lake Michigan. Front. Microbiol. 2015, 6, 1028. [Google Scholar] [CrossRef] [PubMed]
- Mohiuddin, M.M.; Botts, S.R.; Paschos, A.; Schellhorn, H.E. Temporal and spatial changes in bacterial diversity in mixed use watersheds of the Great Lakes region. J. Great Lakes Res. 2019, 45, 109–118. [Google Scholar] [CrossRef]
- Jankowski, K.; Schindler, D.E.; Horner-Devine, M.C. Resource availability and spatial heterogeneity control bacterial community response to nutrient enrichment in lakes. PLoS ONE 2014, 9, e86991. [Google Scholar] [CrossRef] [PubMed]
- Kagalou, I.; Psilovikos, A. Assessment of the typology and the trophic status of two Mediterranean lake ecosystems in Northwestern Greece. Water Resour. 2014, 41, 335–343. [Google Scholar] [CrossRef]
- Papastergiadou, E.; Kagalou, I.; Stefanidis, K.; Retalis, A.; Leonardos, I. Effects of Anthropogenic Influences on the Trophic State, Land Uses and Aquatic Vegetation in a Shallow Mediterranean Lake: Implications for Restoration. Water Resour. Manag. 2010, 24, 415–435. [Google Scholar] [CrossRef]
- Kormas, K.A.; Vardaka, E.; Moustaka-Gouni, M.; Kontoyanni, V.; Petridou, E.; Gkelis, S.; Neofitou, C. Molecular detection of potentially toxic Cyanobacteria and their associated Bacteria in lake water column and sediment. World J. Microbiol. Biotechnol. 2010, 26, 1473–1482. [Google Scholar] [CrossRef]
- Meziti, A.; Tsementzi, D.; Ar Kormas, K.; Karayanni, H.; Konstantinidis, K.T. Anthropogenic effects on bacterial diversity and function along a river-to-estuary gradient in Northwest Greece revealed by metagenomics. Environ. Microbiol. 2016, 18, 4640–4652. [Google Scholar] [CrossRef] [PubMed]
- Meziti, A.; Tsementzi, D.; Rodriguez, R.L.; Hatt, J.K.; Karayanni, H.; Kormas, K.A.; Konstantinidis, K.T. Quantifying the changes in genetic diversity within sequence-discrete bacterial populations across a spatial and temporal riverine gradient. ISME J. 2019, 13, 767–779. [Google Scholar] [CrossRef] [PubMed]
- Baltar, F.; Lindh, M.V.; Parparov, A.; Berman, T.; Pinhassi, J. Prokaryotic community structure and respiration during long-term incubations. Microbiologyopen 2012, 1, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Hobbie, J.E.; Peterson, P.J.; Bettez, N.; Deegan, L.; O’Brien, W.J.; Kling, G.W.; Kipphut, G.W.; Bowden, W.B.; Hershey, A.E. Impact of global change on the biogeochemistry and ecology of an Arctic freshwater system. Polar Res. 1999, 18, 207–214. [Google Scholar] [CrossRef]
- Porter, K.G.; Feig, Y.S. The use of DAPI for identifying and counting aquatic microflora. Limnol. Oceanogr. 1980, 25, 943–948. [Google Scholar] [CrossRef] [Green Version]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glockner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Pruesse, E.Q.C.; Knittel, K.; Fuchs, B.; Ludwig, W.; Peplies, J.; Glöckner, F. Silva: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef] [PubMed]
- Labasque, T.C.C.; Aminot, A.; Kergoat, G. Spectrophotometric Winkler determination of dissolved oxygen: Reexamination of critical factors and reliability. Mar. Chem. 2004, 88, 53–60. [Google Scholar] [CrossRef]
- Apple, J.K.; del Giorgio, P.A.; Kemp, W.M. Temperature regulation of bacterial production, respiration, and growth efficiency in a temperate salt-marsh estuary. Aquat. Microb. Ecol. 2006, 43, 243–254. [Google Scholar] [CrossRef] [Green Version]
- Felip, M.; Pace, M.L.; Cole, J.J. Regulation of planktonic bacterial growth rates: The effects of temperature and resources. Microb. Ecol. 1996, 31, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Berman, T.; Yacobi, Y.Z.; Parparov, A.; Gal, G. Estimation of long-term bacterial respiration and growth efficiency in Lake Kinneret. FEMS Microbiol. Ecol. 2010, 71, 351–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pace, M.L.; Prairie, Y.T. Respiration in Lakes; Oxford University Press: Oxford, UK, 2005. [Google Scholar]
- García-Martín, E.-E.; Serret, P.; Érez-Lorenzo, M. Testing potential bias in marine plankton respiration rates by dark bottle incubations in the NW Iberian shelf: Incubation time and bottle volume. Cont. Shelf Res. 2010, 31, 496–506. [Google Scholar] [CrossRef]
- Pratt, D.M.; Berkson, H. Two sources of error in the oxygen light and dark bottle method. Limnol. Oceanogr. 1954, 4, 328–334. [Google Scholar] [CrossRef]
- Ramette, A. Multivariate analyses in microbial ecology. FEMS Microbiol. Ecol. 2007, 62, 142–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kagalou, I.; Papastergiadou, E.; Leonardos, I. Long term changes in the eutrophication process in a shallow Mediterranean lake ecosystem of W. Greece: Response after the reduction of external load. J. Environ. Manag. 2008, 87, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Newton, R.J.; Shade, A. Lifestyles of rarity: Understanding heterotrophic strategies to inform the ecology of the microbial rare biosphere. Aquat. Microb. Ecol. 2016, 78, 51–63. [Google Scholar] [CrossRef]
- Yang, C.; Wang, Q.; Simon, P.N.; Liu, J.; Liu, L.; Dai, X.; Zhang, X.; Kuang, J.; Igarashi, Y.; Pan, X.; et al. Distinct Network Interactions in Particle-Associated and Free-Living Bacterial Communities during a Microcystis aeruginosa Bloom in a Plateau Lake. Front. Microbiol. 2017, 8, 1202. [Google Scholar] [CrossRef] [PubMed]
- Ghai, R.; Mizuno, C.M.; Picazo, A.; Camacho, A.; Rodriguez-Valera, F. Key roles for freshwater Actinobacteria revealed by deep metagenomic sequencing. Mol. Ecol. 2014, 23, 6073–6090. [Google Scholar] [CrossRef] [PubMed]
- Vardaka, E.; Moustaka-Gouni, M.; Cook, C.M.; Lanaras, T. Cyanobacterial blooms and water quality in Greek waterbodies. J. Appl. Phycol. 2005, 17, 391–401. [Google Scholar] [CrossRef]
- Parulekar, N.N.; Kolekar, P.; Jenkins, A.; Kleiven, S.; Utkilen, H.; Johansen, A.; Sawant, S.; Kulkarni-Kale, U.; Kale, M.; Saebo, M. Characterization of bacterial community associated with phytoplankton bloom in a eutrophic lake in South Norway using 16S rRNA gene amplicon sequence analysis. PLoS ONE 2017, 12, e0173408. [Google Scholar] [CrossRef] [PubMed]
- McManus, G.B.; Kantz, L.A. Molecular and morphological methods for identifying plankton: What makes a successful marriage? J. Plankton Res. 2009, 31, 1119–1129. [Google Scholar] [CrossRef]
- Oikonomou, A.; Katsiapi, M.; Karayanni, H.; Moustaka-Gouni, M.; Kormas, K.A. Plankton microorganisms coinciding with two consecutive mass fish kills in a newly reconstructed lake. Sci. World J. 2012, 2012, 504135. [Google Scholar] [CrossRef] [PubMed]
- Chiang, E.; Schmidt, M.L.; Berry, M.A.; Biddanda, B.A.; Burtner, A.; Johengen, T.H.; Palladino, D.; Denef, V.J. Verrucomicrobia are prevalent in north-temperate freshwater lakes and display class-level preferences between lake habitats. PLoS ONE 2018, 13, e0195112. [Google Scholar]
- Cai, H.; Jiang, H.; Krumholz, L.R.; Yang, Z. Bacterial community composition of size-fractioned aggregates within the phycosphere of cyanobacterial blooms in a eutrophic freshwater lake. PLoS ONE 2014, 9, e102879. [Google Scholar] [CrossRef] [PubMed]
- Cardman, Z.; Arnosti, C.; Durbin, A.; Ziervogel, K.; Cox, C.; Steen, A.D.; Teske, A. Verrucomicrobia are candidates for polysaccharide-degrading bacterioplankton in an arctic fjord of Svalbard. Appl. Environ. Microbiol. 2014, 80, 3749–3756. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Garcia, M.; Brazel, D.M.; Swan, B.K.; Arnosti, C.; Chain, P.S.; Reitenga, K.G.; Xie, G.; Poulton, N.J.; Lluesma Gomez, M.; Masland, D.E.; et al. Capturing single cell genomes of active polysaccharide degraders: An unexpected contribution of Verrucomicrobia. PLoS ONE 2012, 7, e35314. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.; Chen, L.; Liu, Y.; Tao, W.; Zhang, Z.; Liu, H.; Tang, Y. Planktonic and sedimentary bacterial diversity of Lake Sayram in summer. MicrobiologyOpen 2015, 4, 814–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newton, R.J.; Jones, S.E.; Eiler, A.; McMahon, K.D.; Bertilsson, S. A Guide to the Natural History of Freshwater Lake Bacteria. Microbiol. Mol. Boil. Rev. 2011, 75, 14–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiaffino, M.R.; Sánchez, M.L.; Gerea, M.; Unrein, F.; Balagué, V.; Gasol, J.M.; Izaguirre, I. Distribution patterns of the abundance of major bacterial and archaeal groups in Patagonian lakes. J. Plankton Res. 2016, 38, 64–82. [Google Scholar] [CrossRef]
- Doronina, N.; Kaparullina, E.; Trotsenko, Y. The Family Methylophilaceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 869–880. [Google Scholar]
- Salcher, M.M.; Neuenschwander, S.M.; Posch, T.; Pernthaler, J. The ecology of pelagic freshwater methylotrophs assessed by a high-resolution monitoring and isolation campaign. ISME J. 2015, 9, 2442–2453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chistoserdova, L. Methylotrophy in a Lake: From Metagenomics to Single-Organism Physiology. Appl. Environ. Microb. 2011, 77, 4705–4711. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Gong, Y.; Zhou, C.; Zhang, J.; Wang, Z.; Ning, K. Comparison and Interpretation of Taxonomical Structure of Bacterial Communities in Two Types of Lakes on Yun-Gui plateau of China. Sci. Rep. 2016, 6, 30616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlson, C.A.; Bates, N.R.; Ducklow, H.W.; Hansell, D.A. Estimation of bacterial respiration and growth efficiency in the Ross Sea, Antarctica. Aquat. Microb. Ecol. 1999, 19, 229–234. [Google Scholar] [CrossRef]
- Del Giorgio, P.A.; Cole, J.J. Bacterial growth efficiency in natural aquatic ecosystems. Annu. Rev. Ecol. Syst. 1998, 29, 503–541. [Google Scholar] [CrossRef]
- Massana, R.; Pedrós-Alio, C.; Casamayor, E.O.; Gasol, J.M. Changes in marine bacterioplankton phylogenetic composition during incubations designed to measure biogeochemically significant parameters. Limnol. Oceanogr. 2001, 46, 1181–1188. [Google Scholar] [CrossRef] [Green Version]
- Cuevas, L.A.; Egge, J.K.; Thingstad, T.F.; Töpper, B. Organic carbon and mineral nutrient limitation of oxygen consumption, bacterial growth and efficiency in the Norwegian Sea. Polar Biol. 2011, 34, 871–882. [Google Scholar] [CrossRef] [Green Version]
- Herlemann, D.P.; Woelk, J.; Labrenz, M.; Jurgens, K. Diversity and abundance of “Pelagibacterales” (SAR11) in the Baltic Sea salinity gradient. Syst. Appl. Microbiol. 2014, 37, 601–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salcher, M.M.; Pernthaler, J.; Posch, T. Seasonal bloom dynamics and ecophysiology of the freshwater sister clade of SAR11 bacteria ‘that rule the waves’ (LD12). ISME J. 2011, 5, 1242–1252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karayanni, H.; Meziti, A.; Spatharis, S.; Genitsaris, S.; Courties, C.; Kormas, K.A. Changes in Microbial (Bacteria and Archaea) Plankton Community Structure after Artificial Dispersal in Grazer-Free Microcosms. Microorganisms 2017, 5, 31. [Google Scholar] [CrossRef] [PubMed]
- Gattuso, J.P.; Peduzzi, S.; Pizay, M.D.; Tonolla, M. Changes in freshwater bacterial community composition during measurements of microbial and community respiration. J. Plankton Res. 2002, 24, 1197–1206. [Google Scholar] [CrossRef] [Green Version]
- Pomeroy, L.; Wiebe, W.; Deibel, D.; Thompson, R.; Rowe, G.; Pakulski, J. Bacterial responses to temperature and substrate concentration during the Newfoundland spring bloom. Mar. Ecol. Prog. Ser. 1991, 75, 146–159. [Google Scholar] [CrossRef]
- Atkin, O.K.; Bruhn, D.; Tjoelker, M.G. Response of Plant Respiration to Changes in Temperature: Mechanisms and Consequences of Variations in Q10 Values and Acclimation; Springer: Dordrecht, The Netherlands, 2005; Volume 18. [Google Scholar]
- Armitage, D.W. Time-variant species pools shape competitive dynamics and biodiversity-ecosystem function relationships. Proc. R. Soc. B Boil. Sci. 2016, 283, 20161437. [Google Scholar] [CrossRef] [PubMed]
- Langenheder, S.; Lindstrom, E.S.; Tranvik, L.J. Weak coupling between community composition and functioning of aquatic bacteria. Limnol. Oceanogr. 2005, 50, 957–967. [Google Scholar] [CrossRef] [Green Version]
- Becker, S.; Richl, P.; Ernst, A. Seasonal and habitat-related distribution pattern of Synechococcus genotypes in Lake Constance. FEMS Microbiol. Ecol. 2007, 62, 64–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karayanni, H.; Macingo, S.C.; Tolis, V.; Alivertis, D. Diversity of Bacteria in Lakes with Different Chlorophyll Content and Investigation of Their Respiratory Activity through a Long-Term Microcosm Experiment. Water 2019, 11, 467. https://doi.org/10.3390/w11030467
Karayanni H, Macingo SC, Tolis V, Alivertis D. Diversity of Bacteria in Lakes with Different Chlorophyll Content and Investigation of Their Respiratory Activity through a Long-Term Microcosm Experiment. Water. 2019; 11(3):467. https://doi.org/10.3390/w11030467
Chicago/Turabian StyleKarayanni, Hera, Sofia C. Macingo, Vasilis Tolis, and Dimitrios Alivertis. 2019. "Diversity of Bacteria in Lakes with Different Chlorophyll Content and Investigation of Their Respiratory Activity through a Long-Term Microcosm Experiment" Water 11, no. 3: 467. https://doi.org/10.3390/w11030467