
Uptake of Biotinylated Spermine in Astrocytes: Effect of Cx43 siRNA, HIV-Tat Protein and Polyamine Transport Inhibitor on Polyamine Uptake

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Preparation of Primary Cortical Astrocyte Cultures
2.3. SDS-PAGE and Western Blotting Analysis
2.4. siRNA Transfection
2.5. HIV-Tat Protein
2.6. Polyamine Transport Inhibitor (PTI)
2.7. Biotinylated Spermine Uptake and Visualization
2.8. Confocal Microscopy
2.9. Statistics
3. Results
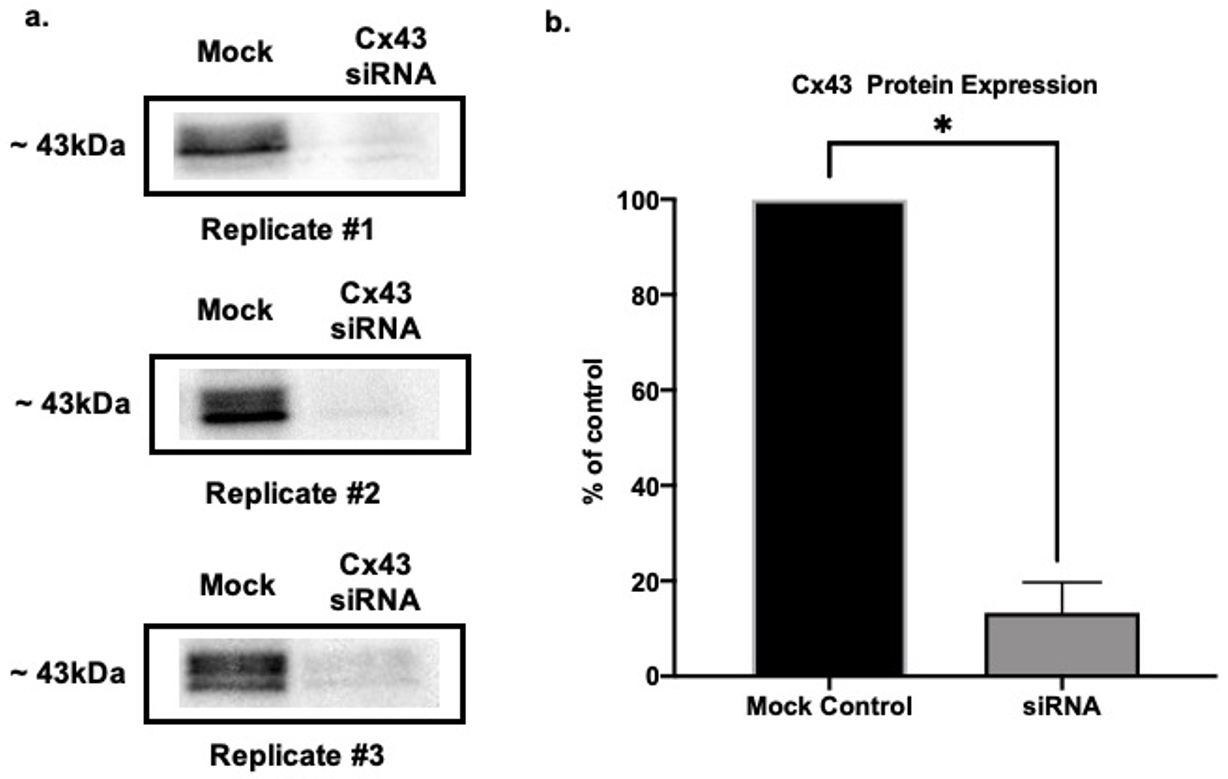
3.1. Cx43 Downregulation Validation via SDS Page and Western Blot
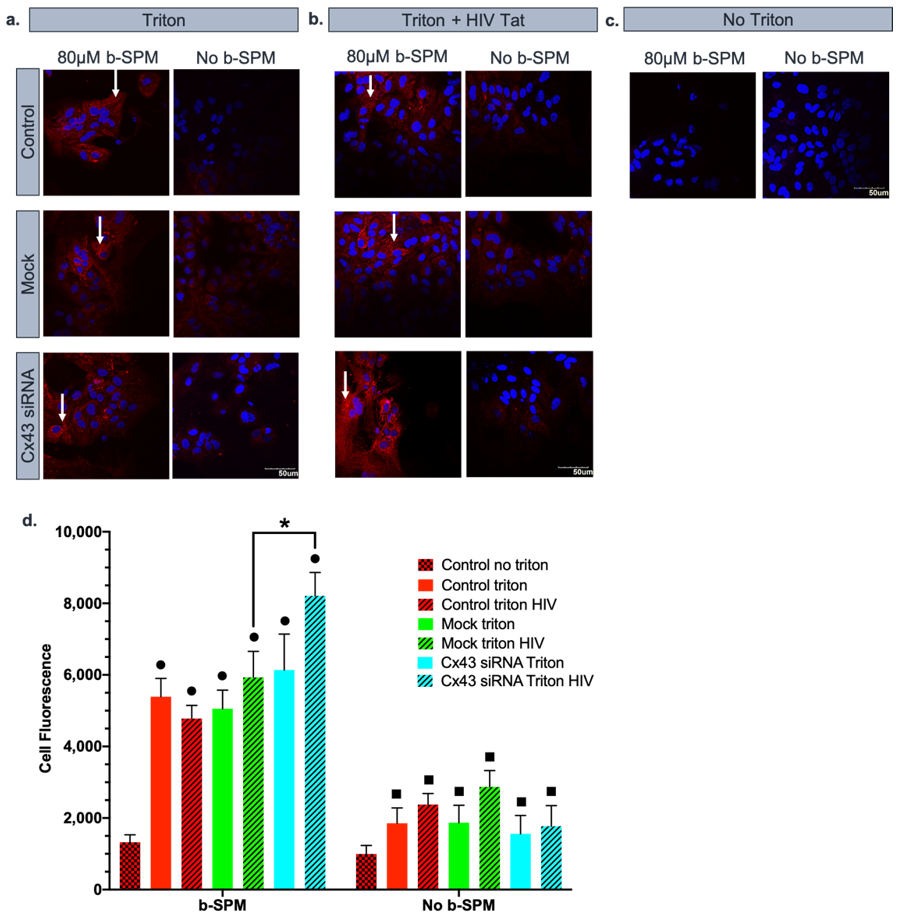
3.2. Contribution of Cx43 Hemichannels to Biotinylated Spermine Uptake into Astrocytes
3.3. Effect of HIV-Tat on the Uptake/Accumulation of b-SPM
3.4. Contribution of Polyamine Transport to Biotinylated Spermine Uptake into Astrocytes
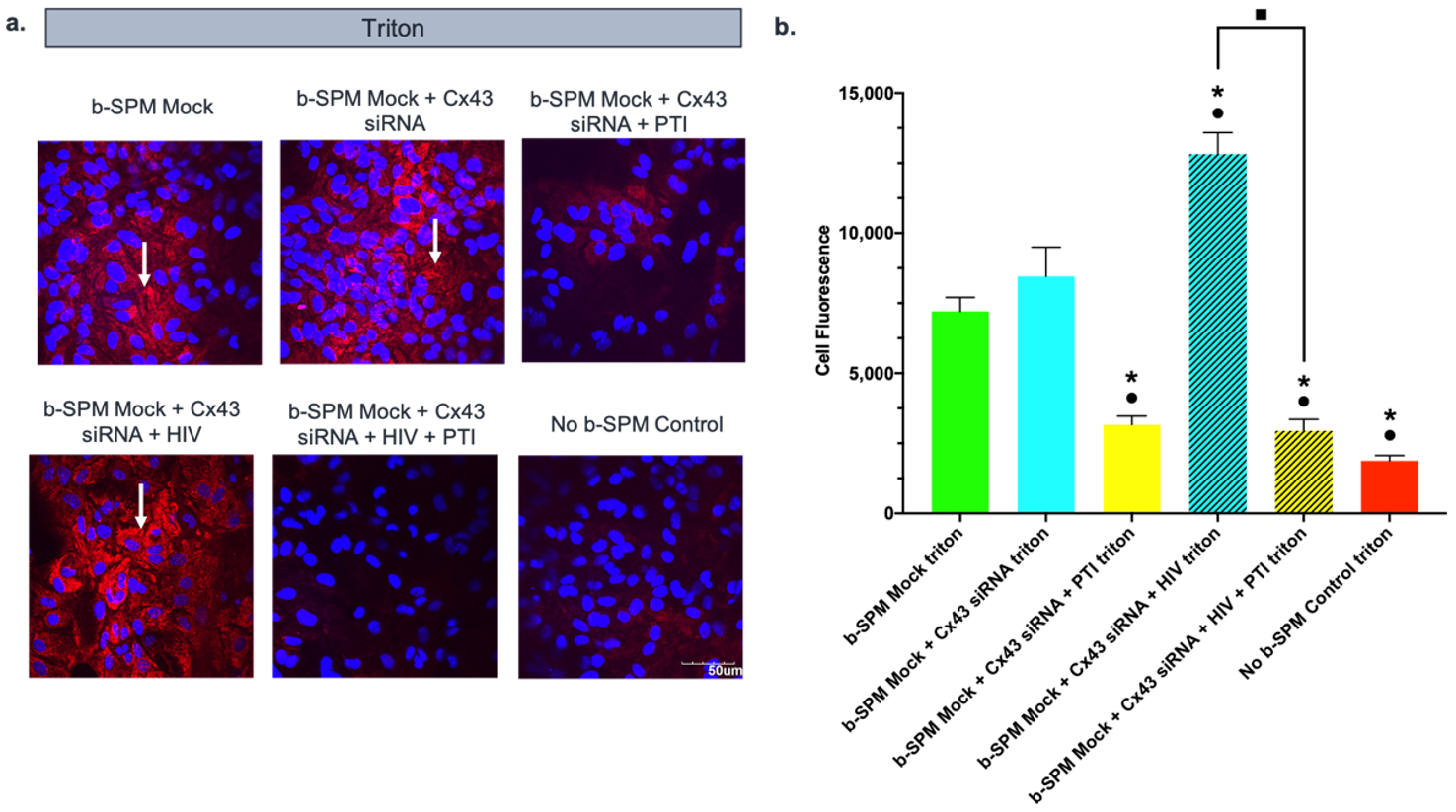
3.5. Effect of PTI on Biotinylated Spermine (b-SPM) Uptake in siRNA Cx43-Treated Astrocytes with and without HIV-Tat Protein
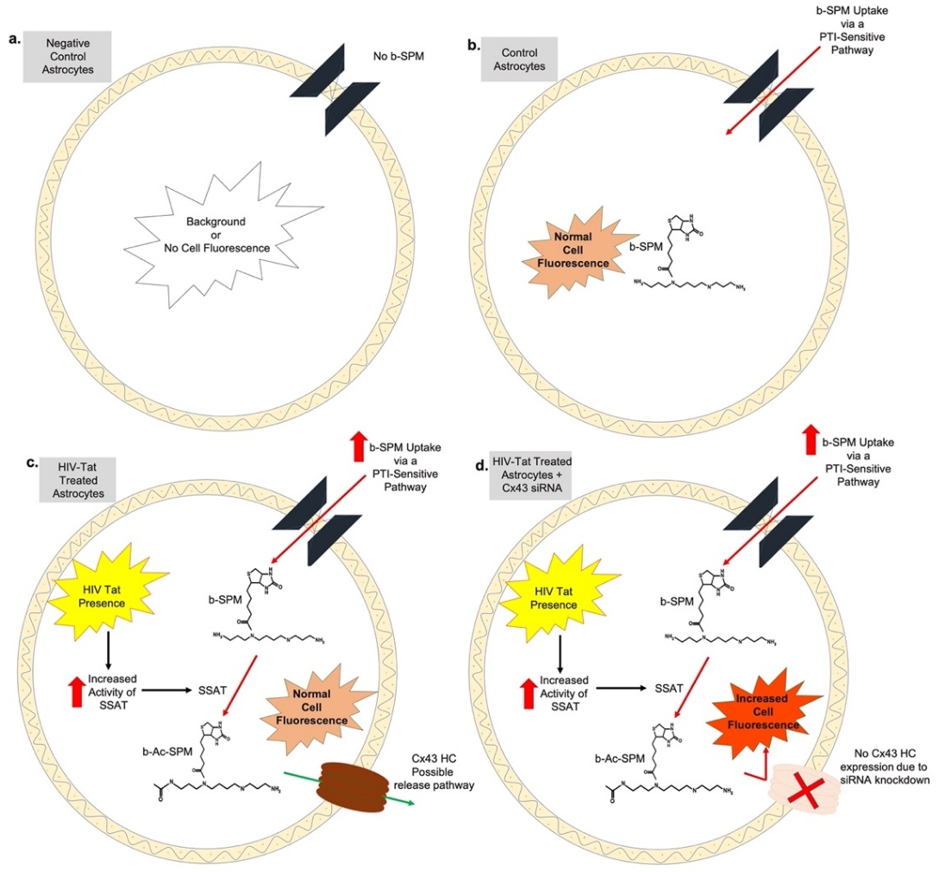
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sharp, P.M. Origins of Human Virus Diversity. Cell 2002, 108, 305–312. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, P.B.; McKeague, I.W.; Eisen, G.; Mullins, C.; Guéye-NDiaye, A.; Mboup, S.; Kanki, P.J. Comparison of HIV-1 and HIV-2 infectivity from a prospective cohort study in Senegal. Stat. Med. 2003, 22, 573–593. [Google Scholar] [CrossRef] [PubMed]
- Spach, D.H.; Hooton, T.M. The HIV Manual: A Guide to Diagnosis and Treatment, 1st ed.; Oxford University Press: New York, NY, USA, 1996. [Google Scholar]
- Doms, R.W.; Trono, D. The plasma membrane as a combat zone in the HIV battlefield. Genes Dev. 2000, 14, 2677–2688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, L.; Roche, M.; Churchill, M.J.; Sterjovski, J.; Ellett, A.; Poumbourios, P.; Sherieff, S.; Sheffief, S.; Wang, B.; Saksena, N.; et al. Tissue-specific sequence alterations in the human immunodeficiency virus type 1 envelope favoring CCR5 usage contribute to persistence of dual-tropic virus in the brain. J. Virol. 2009, 83, 5430–5441. [Google Scholar] [CrossRef] [Green Version]
- Masliah, E.; DeTeresa, R.M.; Mallory, M.E.; Hansen, L.A. Changes in pathological findings at autopsy in AIDS cases for the last 15 years. AIDS 2000, 14, 69–74. [Google Scholar] [CrossRef]
- Zhou, L.; Saksena, N.K. HIV Associated Neurocognitive Disorders. Infect. Dis. Rep. 2013, 5 (Suppl. 1), 38–50. [Google Scholar] [CrossRef] [Green Version]
- Nightingale, S.; Winsto, A.; Letendre, S.; Michael, B.D.; McArthur, J.C.; Khoo, S.; Solomon, T. Controversies in HIV-associated neurocognitive disorders. Lancet Neurol. 2014, 13, 1139–1151. [Google Scholar] [CrossRef] [Green Version]
- Paris, J.J.; Singh, H.D.; Carey, A.N.; McLaughlin, J.P. Exposure to HIV-1 Tat in brain impairs sensorimotor gating and activates microglia in limbic and extralimbic brain regions of male mice. Behav. Brain. Res. 2015, 291, 209–218. [Google Scholar] [CrossRef] [Green Version]
- Jeang, K.T.; Chang, Y.; Berkhout, B.; Hammarskjöld, M.L.; Rekosh, D. Regulation of HIV expression: Mechanisms of action of Tat and Rev. AIDS 1991, 5 (Suppl. 2), 3–14. [Google Scholar] [CrossRef]
- Parada, C.A.; Roeder, R.G. A novel RNA polymerase II-containing complex potentiates Tat-enhanced HIV-1 transcription. EMBO J. 1999, 18, 3688–3701. [Google Scholar] [CrossRef] [Green Version]
- Chandra, T.; Maier, W.; König, H.-G.; Hirzel, K.; Kögel, D.; Schüler, T.; Chandra, A.; Demirhan, I.; Laube, B. Molecular interactions of the type 1 human immunodeficiency virus transregulatory protein Tat with N-methyl-d-aspartate receptor subunits. Neuroscience 2005, 134, 145–153. [Google Scholar] [CrossRef]
- Potter, M.C.; Figuera-Losada, M.; Rojas, C.; Slusher, B.S. Targeting the glutamatergic system for the treatment of HIV-associated neurocognitive disorders. J. Neuroimmune Pharmacol. 2013, 8, 594–607. [Google Scholar] [CrossRef] [Green Version]
- Copeland, K.F.T. Modulation of HIV-1 transcription by cytokines and chemokines. Mini-Rev. Med. Chem. 2005, 5, 1093–1101. [Google Scholar] [CrossRef]
- Royal, W.; Zhang, L.; Guo, M.; Jones, O.; Davis, H.; Bryant, J.L. Immune activation, viral gene product expression and neurotoxicity in the HIV-1 transgenic rat. J. Neuroimmunol. 2012, 247, 16–24. [Google Scholar] [CrossRef] [Green Version]
- Kruman, I.I.; Nath, A.; Mattson, M.P. HIV-1 protein Tat induces apoptosis of hippocampal neurons by a mechanism involving caspase activation, calcium overload, and oxidative stress. Exp. Neurol. 1998, 154, 276–288. [Google Scholar] [CrossRef]
- Krauss, M.; Langnaese, K.; Richter, K.; Brunk, I.; Wieske, M.; Ahnert-Hilger, G.; Veh, R.W.; Laube, G. Spermidine synthase is prominently expressed in the striatal patch compartment and in putative interneurones of the matrix compartment. J. Neurochem. 2006, 97, 174–189. [Google Scholar] [CrossRef]
- Krauss, M.; Weiss, T.; Langnaese, K.; Richter, K.; Kowski, A.; Veh, R.W.; Laube, G. Cellular and subcellular rat brain spermidine synthase expression patterns suggest region-specific roles for polyamines, including cerebellar pre-synaptic function. J. Neurochem. 2007, 103, 679–693. [Google Scholar] [CrossRef]
- Laube, G.; Veh, R.W. Astrocytes, not neurons, show most prominent staining for spermidine/spermine-like immunoreactivity in adult rat brain. GLIA 1997, 19, 171–179. [Google Scholar] [CrossRef]
- Biedermann, B.; Skatchkov, S.N.; Brunk, I.; Bringmann, A.; Pannicke, T.; Bernstein, H.G.; Faude, F.; Germer, A.; Veh, R.; Reichenbach, A. Spermine/spermidine is expressed by retinal glial (Muller) cells and controls distinct K+ channels of their membrane. GLIA 1998, 23, 209–220. [Google Scholar] [CrossRef]
- Skatchkov, S.N.; Eaton, M.J.; Krusek, J.; Veh, R.W.; Biedermann, B.; Bringmann, A.; Pannicke, T.; Orkand, R.K.; Reichenbach, A. Spatial distribution of spermine/spermidine content and K(+)-current rectification in frog retinal glial (Muller) cells. GLIA 2000, 31, 84–90. [Google Scholar] [CrossRef]
- Lambotte, O.; Deiva, K.; Tardieu, M. HIV-1 persistence, viral reservoir, and the central nervous system in the HAART era. Brain Pathol. 2003, 13, 95–103. [Google Scholar] [CrossRef]
- Gorry, P.R.; Ong, C.; Thorpe, J.; Bannwarth, S.; Thompson, K.A.; Gatignol, A.; Vesselingh, S.L.; Purcell, D.F. Astrocyte infection by HIV-1: Mechanisms of restricted virus replication, and role in the pathogenesis of HIV-1-associated dementia. Curr. HIV Res. 2003, 1, 463–473. [Google Scholar] [CrossRef]
- Li, G.H.; Henderson, L.; Nath, A. Astrocytes as an HIV Reservoir: Mechanism of HIV Infection. Curr. HIV Res. 2016, 14, 373–381. [Google Scholar] [CrossRef]
- Mounce, B.C.; Poirier, E.Z.; Passoni, G.; Simon-Loriere, E.; Cesaro, T.; Prot, M.; Stapleford, K.A.; Moratorio, G.; Sakuntabhai, A.; Levraud, J.P.; et al. Interferon-Induced Spermidine-Spermine Acetyltransferase and Polyamine Depletion Restrict Zika and Chikungunya Viruses. Cell Host Microbe 2016, 20, 167–177. [Google Scholar] [CrossRef] [Green Version]
- Frankel, E.A.; Bevilacqua, P.C.; Keating, C.D. Polyamine/Nucleotide Coacervates Provide Strong Compartmentalization of Mg2+, Nucleotides, and RNA. Langmuir 2016, 32, 2041–2049. [Google Scholar] [CrossRef]
- Merali, S.; Barrero, C.A.; Sacktor, N.C.; Haughey, N.J.; Datta, P.K.; Langford, D.; Khalili, K. Polyamines: Predictive Biomarker for HIV-Associated Neurocognitive Disorders. J. AIDS Res. Treat. 2014, 5, 1000312. [Google Scholar] [CrossRef]
- Skatchkov, S.N.; Woodbury-Fariña, M.A.; Eaton, M. The Role of Glia in Stress. Psychiatr. Clin. N. Am. 2014, 37, 653–678. [Google Scholar] [CrossRef] [Green Version]
- Skatchkov, S.N.; Antonov, S.M.; Eaton, M.J. Glia and glial polyamines. Role in brain function in health and disease. Biochemistry 2016, 10, 73–98. [Google Scholar] [CrossRef]
- Gilad, G.M.; Gilad, V.H. Polyamine uptake, binding and release in rat brain. Eur. J. Pharmacol. 1991, 193, 41–46. [Google Scholar] [CrossRef]
- Ingoglia, N.A.; Sharma, S.C.; Pilchman, J.; Baranowski, K.; Sturman, J.A. Axonal transport and transcellular transfer of nucleosides and polyamines in intact and regenerating optic nerves of goldfish: Speculation on the axonal regulation of periaxonal cell metabolism. J. Neurosci. 1982, 2, 1412–1423. [Google Scholar] [CrossRef]
- Olsen, M.L.; Khakh, B.S.; Skatchkov, S.N.; Zhou, M.; Lee, C.J.; Rouach, N. New Insights on Astrocyte Ion Channels: Critical for Homeostasis and Neuron-Glia Signaling. J. Neurosci. 2015, 35, 13827–13835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malpica-Nieves, C.J.; Rivera-Aponte, D.E.; Tejeda-Bayron, F.A.; Mayor, A.M.; Phanstiel, O.; Veh, R.W.; Eaton, M.J.; Skatchkov, S.N. The involvement of polyamine uptake and synthesis pathways in the proliferation of neonatal astrocytes. Amino Acids 2020, 52, 1169–1180. [Google Scholar] [CrossRef] [PubMed]
- Lent, R.; Azevedo, F.A.; Andrade-Moraes, C.H.; Pinto, A.V. How many neurons do you have? Some dogmas of quantitative neuroscience under revision. Eur. J. Neurosci. 2012, 35, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, T.; Knauer, H.; Schauer, A.; Buttner, S.; Ruckenstuhl, C.; Carmona-Gutierrez, D.; Ring, J.; Schroeder, S.; Magnes, C.; Antonacci, L.; et al. Induction of autophagy by spermidine promotes longevity. Nat. Cell Biol. 2009, 11, 1305–1314. [Google Scholar] [CrossRef]
- Viltard, M.; Durand, S.; Pérez-Lanzón, M.; Aprahamian, F.; Lefevre, D.; Leroy, C.; Madeo, F.; Kroemer, G.; Friedlander, G. The metabolomic signature of extreme longevity: Naked mole rats versus mice. Aging 2019, 11, 4783–4800. [Google Scholar] [CrossRef]
- Benedikt, J.; Inyushin, M.; Kucheryavykh, Y.V.; Rivera, Y.; Kucheryavykh, L.Y.; Nichols, C.G.; Eaton, M.J.; Skatchkov, S.N. Intracellular polyamines enhance astrocytic coupling. Neuroreport 2012, 23, 1021–1025. [Google Scholar] [CrossRef]
- Kucheryavykh, L.Y.; Benedikt, J.; Cubano, L.A.; Skatchkov, S.N.; Bukauskas, F.F.; Kucheryavykh, Y.V. Polyamines preserve connexin 43-mediated gap junctional communication during intracellular hypercalcemia and acidosis. Neuroreport 2017, 28, 208–213. [Google Scholar] [CrossRef] [Green Version]
- Skatchkov, S.N.; Bukauskas, F.F.; Benedikt, J.; Inyushin, M.; Kucheryavykh, Y.V. Intracellular spermine prevents acid-induced uncoupling of Cx43 gap junction channels. Neuroreport 2015, 26, 528–532. [Google Scholar] [CrossRef]
- Li, D.C.; Nichols, C.G.; Sala-Rabanal, M. Role of a Hydrophobic Pocket in Polyamine Interactions with the Polyspecific Organic Cation Transporter OCT3. J. Biol. Chem. 2015, 290, 27633–27643. [Google Scholar] [CrossRef] [Green Version]
- Makarov, V.; Kucheryavykh, L.; Kucheryavykh, Y.; Rivera, A.; Eaton, M.J.; Skatchkov, S.N.; Inyushin, M. Transport Reversal during Heteroexchange: A Kinetic Study. J. Biophys. 2013, 2013, 683256. [Google Scholar] [CrossRef] [Green Version]
- Sala-Rabanal, M.; Li, D.C.; Dake, G.R.; Kurata, H.T.; Inyushin, M.; Skatchkov, S.N.; Nichols, C.G. Polyamine transport by the polyspecific organic cation transporters OCT1, OCT2, and OCT3. Mol. Pharm. 2013, 10, 1450–1458. [Google Scholar] [CrossRef] [Green Version]
- Inazu, M.; Takeda, H.; Matsumiya, T. Expression and functional characterization of the extraneuronal monoamine transporter in normal human astrocytes. J. Neurochem. 2003, 84, 43–52. [Google Scholar] [CrossRef]
- Gitto, S.B.; Pandey, V.; Oyer, J.L.; Copik, A.J.; Hogan, F.C.; Phanstiel, O.; Altomare, D.A. Difluoromethylornithine Combined with a Polyamine Transport Inhibitor Is Effective against Gemcitabine Resistant Pancreatic Cancer. Mol. Pharm. 2018, 15, 369–376. [Google Scholar] [CrossRef]
- Muth, A.; Madan, M.; Archer, J.J.; Ocampo, N.; Rodriguez, L.; Phanstiel, O. Polyamine Transport Inhibitors: Design, Synthesis, and Combination Therapies with Difluoromethylornithine. J. Med. Chem. 2014, 57, 348–363. [Google Scholar] [CrossRef]
- Hopman, A.H.; Ramaekers, F.C.; Speel, E.J. Rapid synthesis of biotin-, digoxigenin-, trinitrophenyl-, and fluorochrome-labeled tyramides and their application for In situ hybridization using CARD amplification. J. Histochem. Cytochem. 1998, 46, 771–777. [Google Scholar] [CrossRef] [Green Version]
- Madai, V.I.; Poller, W.C.; Peters, D.; Berger, J.; Paliege, K.; Bernard, R.; Veh, R.W.; Laube, G. Synaptic localisation of agmatinase in rat cerebral cortex revealed by virtual pre-embedding. Amino Acids 2012, 43, 1399–1403. [Google Scholar] [CrossRef]
- Hammond, L. Measuring Cell Fluorescence Using ImageJ—The Open Lab Book v1.0. The Open Lab Book. 2014. Available online: https://theolb.readthedocs.io/en/latest/imaging/measuring-cell-fluorescence-using-imagej.html (accessed on 11 August 2012).
- Livnah, O.; Bayer, E.A.; Wilchek, M.; Sussman, J.L. Three-dimensional structures of avidin and the avidin-biotin complex. Proc. Natl. Acad. Sci. USA 1993, 90, 5076–5080. [Google Scholar] [CrossRef] [Green Version]
- Rosano, C.; Arosio, P.; Bolognesi, M. The X-ray three-dimensional structure of avidin. Biomed. Eng. 1999, 16, 5–12. [Google Scholar] [CrossRef]
- Mani, K.; Sandgren, S.; Lilja, J.; Cheng, F.; Svensson, K.; Persson, L.; Belting, M. HIV-Tat protein transduction domain specifically attenuates growth of polyamine deprived tumor cells. Mol. Cancer Ther. 2007, 6, 782–788. [Google Scholar] [CrossRef] [Green Version]
- Minois, N.; Carmona-Gutierrez, D.; Madeo, F. Polyamines in aging and disease. Aging 2011, 3, 716–732. [Google Scholar] [CrossRef] [Green Version]
- Wallace, H.M.; Fraser, A.V. Inhibitors of polyamine metabolism: Review article. Amino Acids 2004, 26. [Google Scholar] [CrossRef]
- Bernstein, H.G.; Muller, M. The cellular localization of the L-ornithine decarboxylase/polyamine system in normal and diseased central nervous systems. Prog. Neurobiol. 1999, 57, 485–505. [Google Scholar] [CrossRef]
- Madan, M.; Patel, A.; Skruber, K.; Geerts, D.; Altomare, D.A.; Phanstiel, O. ATP13A3 and caveolin-1 as potential biomarkers for difluoromethylornithine-based therapies in pancreatic cancers. Am. J. Cancer Res. 2016, 6, 1231–1252. [Google Scholar]
- Busch, A.E.; Quester, S.; Ulzheimer, J.C.; Waldegger, S.; Gorboulev, V.; Arndt, P.; Lang, F.; Koepsell, H. Electrogenic properties and substrate specificity of the polyspecific rat cation transporter rOCT1. J. Biol. Chem. 1996, 271, 32599–32604. [Google Scholar] [CrossRef] [Green Version]
- Slitt, A.L.; Cherrington, N.J.; Hartley, D.P.; Leazer, T.M.; Klaassen, C.D. Tissue distribution and renal developmental changes in rat organic cation transporter mRNA levels. Drug Metab. Dispos. 2002, 30, 212–219. [Google Scholar] [CrossRef] [Green Version]
- Cui, M.; Aras, R.; Christian, W.V.; Rappold, P.M.; Hatwar, M.; Panza, J.; Jackson-Lewis, V.; Javitch, J.A.; Ballatori, N.; Przedborski, S.; et al. The organic cation transporter-3 is a pivotal modulator of neurodegeneration in the nigrostriatal dopaminergic pathway. Proc. Natl. Acad. Sci. USA 2009, 106, 8043–8048. [Google Scholar] [CrossRef] [Green Version]
- Dot, J.; Lluch, M.; Blanco, I.; Rodríguez-Alvarez, J. Polyamine uptake in cultured astrocytes: Characterization and modulation by protein kinases. J. Neurochem. 2000, 75, 1917–1926. [Google Scholar] [CrossRef]
- Gründemann, D.; Koster, S.; Kiefer, N.; Breidert, T.; Engelhardt, M.; Spitzenberger, F.; Obermuller, N.; Schomig, E. Transport of monoamine transmitters by the organic cation transporter type 2, OCT2. J. Biol. Chem. 1998, 273, 30915–30920. [Google Scholar] [CrossRef] [Green Version]
- Gründemann, D.; Liebich, G.; Kiefer, N.; Köster, S.; Schömig, E. Selective substrates for non-neuronal monoamine transporters. Mol. Pharmacol. 1999, 56, 1–10. [Google Scholar] [CrossRef]
- Schömig, E.; Lazar, A.; Gründemann, D. Extraneuronal monoamine transporter and organic cation transporters 1 and 2: A review of transport efficiency. In Neurotransmitter Transporters; Springer: Berlin/Heidelberg, Germany, 2006. [Google Scholar] [CrossRef]
- Spray, D.C.; Ye, Z.C.; Ransom, B.R. Functional connexin “hemichannels”: A critical appraisal. GLIA 2006, 54, 758–773. [Google Scholar] [CrossRef]
- Ye, Z.C.; Wyeth, M.S.; Baltan-Tekkok, S.; Ransom, B.R. Functional hemichannels in astrocytes: A novel mechanism of glutamate release. J. Neurosci. 2003, 23, 3588–3596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kucheryavykh, Y.V.; Inyushin, M.Y.; Maldonado, H.M.; Burnashev, N.; Nichols, C.G.; Eaton, M.J.; Skatchkov, S.N. Spermine potentiates hemichannel opening in cortical astrocytes. In Proceedings of the 35rd Annual Meeting of the Society for Neuroscience, Washington, DC, USA, 13–16 November 2005. [Google Scholar]
- Cohen, J.E.; Fields, R.D. Extracellular Calcium Depletion in Synaptic Transmission. Neuroscientist 2004, 10, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, G.S.; Moreno, A.P.; Bechberger, J.F.; Hearn, S.S.; Shivers, R.R.; MacPhee, D.J.; Zhang, Y.C.; Naus, C.C.G. Evidence That Disruption of Connexon Particle Arrangements in Gap Junction Plaques Is Associated with Inhibition of Gap Junctional Communication by a Glycyrrhetinic Acid Derivative. Exp. Cell Res. 1996, 222, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Willebrords, J.; Maes, M.; Crespo Yanguas, S.; Vinken, M. Inhibitors of connexin and pannexin channels as potential therapeutics. Pharmacol. Ther. 2017, 180, 144–160. [Google Scholar] [CrossRef] [Green Version]
- Charveriat, M.; Mouthon, F.; Rein, W.; Verkhratsky, A. Connexins as therapeutic targets in neurological and neuropsychiatric disorders. Biochim. Biophys. Acta Mol. Basis Dis. 2021, 1867, 166098. [Google Scholar] [CrossRef]
- Massmann, V.; Edemir, B.; Schlatter, E.; Al-Monajjed, R.; Harrach, S.; Klassen, P.; Holle, S.K.; Sindic, A.; Dobrivojevic, M.; Pavenstädt, H.; et al. The organic cation transporter 3 (OCT3) as molecular target of psychotropic drugs: Transport characteristics and acute regulation of cloned murine OCT3. Pflugers Arch. 2014, 466, 517–527. [Google Scholar] [CrossRef]
- Koepsell, H. Organic Cation Transporters in Health and Disease. Pharmacol. Rev. 2020, 72, 253–319. [Google Scholar] [CrossRef]
- Poulin, R.; Casero, R.A.; Soulet, D. Recent advances in the molecular biology of metazoan polyamine transport. Amino Acids 2012, 42, 711–723. [Google Scholar] [CrossRef] [Green Version]
- Berman, J.W.; Carvallo, L.; Buckner, C.M.; Luers, A.; Prevedel, L.; Bennett, M.V.; Eugenin, E.A. HIV-tat alters Connexin43 expression and trafficking in human astrocytes: Role in NeuroAIDS. J. Neuroinflam. 2016, 13, 54. [Google Scholar] [CrossRef] [Green Version]
- Ray, R.M.; Bhattacharya, S.; Bavaria, M.N.; Viar, M.J.; Johnson, L.R. Spermidine, a sensor for antizyme 1 expression regulates intracellular polyamine homeostasis. Amino Acids 2014, 46, 2005–2013. [Google Scholar] [CrossRef] [Green Version]
- Vialou, V.; Balasse, L.; Callebert, J.; Launay, J.M.; Giros, B.; Gautron, S. Altered aminergic neurotransmission in the brain of organic cation transporter 3-deficient mice. J. Neurochem. 2008, 106, 1471–1482. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malpica-Nieves, C.J.; Rivera, Y.; Rivera-Aponte, D.E.; Phanstiel, O.; Veh, R.W.; Eaton, M.J.; Skatchkov, S.N. Uptake of Biotinylated Spermine in Astrocytes: Effect of Cx43 siRNA, HIV-Tat Protein and Polyamine Transport Inhibitor on Polyamine Uptake. Biomolecules 2021, 11, 1187. https://doi.org/10.3390/biom11081187
Malpica-Nieves CJ, Rivera Y, Rivera-Aponte DE, Phanstiel O, Veh RW, Eaton MJ, Skatchkov SN. Uptake of Biotinylated Spermine in Astrocytes: Effect of Cx43 siRNA, HIV-Tat Protein and Polyamine Transport Inhibitor on Polyamine Uptake. Biomolecules. 2021; 11(8):1187. https://doi.org/10.3390/biom11081187
Chicago/Turabian StyleMalpica-Nieves, Christian J., Yomarie Rivera, David E. Rivera-Aponte, Otto Phanstiel, Rüdiger W. Veh, Misty J. Eaton, and Serguei N. Skatchkov. 2021. "Uptake of Biotinylated Spermine in Astrocytes: Effect of Cx43 siRNA, HIV-Tat Protein and Polyamine Transport Inhibitor on Polyamine Uptake" Biomolecules 11, no. 8: 1187. https://doi.org/10.3390/biom11081187