Meiofaunal Dynamics and Heterogeneity along Salinity and Trophic Gradients in a Mediterranean Transitional System


1
DiSB, Università degli Studi di Urbino Carlo Bo, Campus Scientifico Enrico Mattei, Località Crocicchia, 61029 Urbino, Italy
2
Dipartimento di Biologia, Università di Roma Tor Vergata, Via della Ricerca Scientifica, 1, 00133 Roma, Italy
3
Consorzio Nazionale Interuniversitario per le Scienze del Mare (CoNISMa), Piazzale Flaminio 9, 00196 Rome, Italy
4
Consiglio Nazionale delle Ricerche, Istituto per lo studio degli impatti Antropici e Sostenibilità in ambiente marino (CNR-IAS), Località Sa Mardini, Torregrande, 09170 Oristano, Italy
*
Authors to whom correspondence should be addressed.
Water 2019, 11(7), 1488; https://doi.org/10.3390/w11071488
Submission received: 10 May 2019
/
Revised: 1 July 2019
/
Accepted: 13 July 2019
/
Published: 18 July 2019
(This article belongs to the Special Issue Ecological Status Assessment of Transitional Waters)
Abstract
:The spatiotemporal variation in meiofaunal assemblages were investigated for the first time in the Cabras Lagoon, the largest transitional system in the Sardinian Island (W-Mediterranean Sea). Two main environmental (salinity and trophic) gradients highlighted a significant separation of the three study sites across the lagoon, which were consistent through time. The environmental variability and habitat heterogeneity of the Cabras Lagoon influenced the meiofauna. In particular, salinity and dissolved oxygen, primarily, shaped the meiofaunal assemblage structure at the seaward site which was significantly different from both the riverine and the organically enriched sites. On the other hand, the trophic components (e.g., organic matter, Chlorophyll-a, and phaeopigments) and the different degrees of confinement and saprobity among sites were the secondary factors contributing mostly to the separation between the latter two sites. The lack of significant differences in the temporal comparison of the meiofaunal assemblage structure along with the very low contribution of temperature to the meiofaunal ordination indicated that this assemblage was more affected by spatial rather than by temporal variation. This pattern was also supported by significant differences between the three sites in several univariate measures, including total number of individuals, number of taxa, Pielou’s evenness, and the ratio between nematodes and copepods. Thus, the present study corroborates the hypothesis that meiofaunal organisms are good indicators of the spatial heterogeneity in transitional waters (TWs) and could have a greater species richness than that expected. Indeed, the Cabras Lagoon overall showed one of the highest meiofaunal richness values found from both Mediterranean and European TWs.
1. Introduction
Transitional waters, being a continuum between continental and marine ecosystems, represent areas with high environmental heterogeneity. As such, there is a complex association between abiotic and biotic components that makes these water bodies ideal to study the distribution and dynamics of the benthic assemblages with the aim to further our understanding of the ecosystem functioning [1,2]. Lagoons have a historically relevant “social” value because they offer a high biological productivity [3]. For this reason, they host many human activities (i.e., fisheries, aquaculture, agriculture, industry, and tourism [4]) that, on the other side, have endangered their integrity and ecological quality status as well [5,6]. Therefore, there is general agreement among the scientific community, which is also recognized by legislations worldwide (e.g., the US Clean Water Act, European Water Framework Directive, Marine Strategy Framework Directive, and the National Water Act in South Africa), about the need to assess their health status and ensure proper management of their resources [4,7].
Meiofauna are small benthic invertebrates that have a well-recognized role in the food webs of lagoon systems connecting microbial components to higher trophic levels that contributes to the overall carbon fluxes and organic matter mineralization [8,9]. Because of their high taxonomic diversity, rapid generation times, lack of larval stages, and various life strategies meiofaunal organisms are considered excellent bioindicators of natural or anthropogenic stressful conditions [10,11,12]. However, their role in ecosystems tends to be overlooked, mainly due to the lack of taxonomists and the small size of meiofauna, which require time and the appropriate techniques for their study [13].
In the Mediterranean basin, there are more than 100 coastal lagoons, half of which have available physico-chemical or ecological data in the scientific literature [4]. Among them, the largest amount of information on meiofaunal spatial pattern is available for the upper Adriatic Sea, including the Venice lagoon [14,15,16,17,18,19,20]. Meiofaunal diversity and assemblage structure are also well-documented in the southern part of the Adriatic Sea, including the Lesina and Varano lagoons [6,8,17,21,22,23]. Instead, meiofaunal studies in transitional environments along the Tyrrhenian coast, with the exception of the Stagnone of Marsala (Western Sicily), are largely lacking [17,24]. Furthermore, most of the available literature on meiofauna from coastal lagoons takes into consideration the spatial pattern of the assemblages, while only in a few cases their temporal dynamics is reported [2,8,24,25,26]. Finally, little is known on spatiotemporal dynamics of meiofauna in Mediterranean transitional systems characterized by different physico-chemical gradients related to the riverine inflow, the connection to the sea, and the organic matter (OM) enrichment of sediments.
Within the Tyrrhenian coast, the Sardinian Island is one of the richest Italian regions in number and extension of lagoons [27], yet knowledge on meiofaunal composition and distribution in these systems is absent. In the present study, we describe for the first time the spatiotemporal variation in meiofaunal assemblages in the Cabras Lagoon, the largest and most complex transitional system in the Sardinia Island. This lagoon is characterized by a large environmental heterogeneity, with an increasing salinity along its main longitudinal axis and varying degrees of trophic condition across the basin [28,29,30]. For these reasons, it represents a valuable case-study in which to test the general hypotheses on the meiofaunal dynamics in these highly variable systems. Our main objectives were to investigate the pattern of spatial variation in meiofaunal diversity and community structure in relation to the main environmental gradients, and to assess whether this pattern was consistent through time. In particular, we tested whether spatiotemporal variation could be identified in: (1) the whole meiofaunal assemblage of the three sites in terms of (i) total number of individuals, (ii) total number of taxa, and (iii) Shannon diversity (H’) and Pielou’s evenness (J) indices; and (2) the abundance of dominant taxa, including the ratio between nematodes and copepods. We anticipate that the response of meiofaunal assemblages to the environmental drivers (both in water and sediments) identified in the present study will provide one of the few evidences of the importance of meiofaunal studies to further our understanding of the functioning of Mediterranean lagoons.
2. Materials and Methods
2.1. Study Area and Sampling Sites
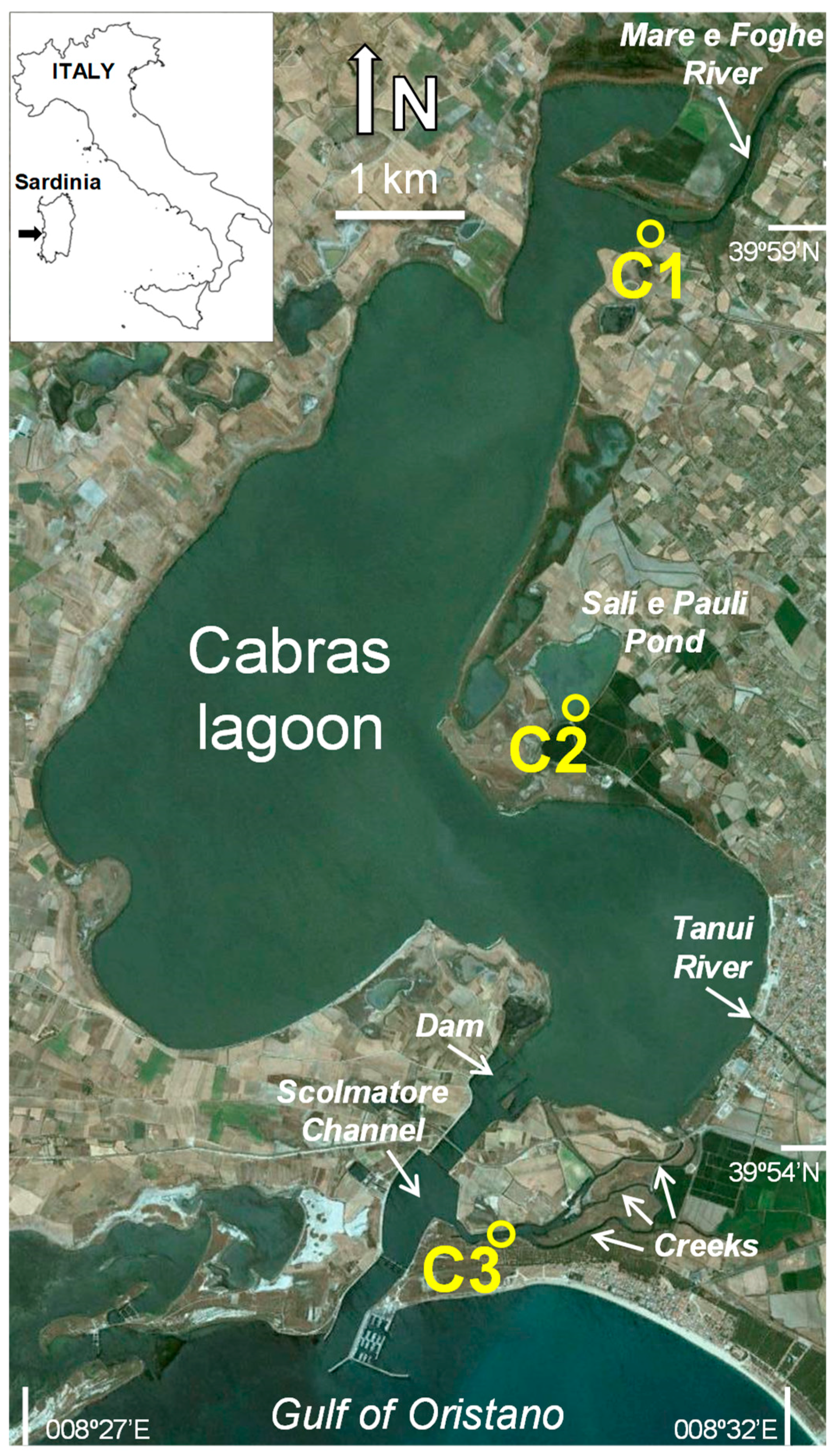
The Cabras Lagoon (central-western Sardinia; Figure 1) is the largest lagoon in the Sardinia island, with a surface area of 22 km2 and a watershed of ~430 km2 inhabited by approximately 38,000 people. Its main freshwater riverine source is the Rio Mare e Foghe located in the northern sector of the lagoon, with a minor contribution from the Rio Tanui, southward. The lagoon is connected to the adjacent Gulf of Oristano only via three narrow creeks that flow into a large channel (“scolmatore”) built in the late 70’s, closed in proximity of the lagoon by a 30 cm high dam. In the last two decades, the Cabras Lagoon has been extensively investigated from various perspectives and using different approaches, including physical/modeling [31,32], biogeochemical [33,34], biological [35,36,37,38], and ecological [5,39,40]. However, while several studies have been conducted in the Cabras Lagoon on the macrozoobenthos [28,29,30,41,42], nothing is known regarding the spatiotemporal variation in meiofaunal assemblages. In fact, no such studies are available for transitional waters in the Sardinian Island, one of the richest Italian regions in number and extension of lagoons [27], with only few examples conducted in fully marine coastal waters [43,44].
For the present study, three sites (C1, C2, and C3; Figure 1) were selected along the longitudinal axis of the Cabras Lagoon, being representative of different environmental (e.g., salinity, confinement, and sediment grain-size) and trophic (e.g., sediment OM and phytopigments) conditions. Site C1 was located in the northern sector of the lagoon, connected to the main freshwater tributary the Rio Mare e Foghe. This site was characterized by sandy sediments, low OM content of sediments, and the presence of halophytic vegetation (Phragmites sp.) along the shore. Site C2, was located in the satellite pond of Sali e Pauli and surrounded by halophytic vegetation (Salicornia sp.). This site was highly confined and characterized by a high OM content of sediments [33]. Biofilm-forming cyanobacterial strains with extremely growth rates were also found here [45,46]. Site C3 was located in the southern sector of the lagoon, at the confluence of the three creeks connecting the Cabras Lagoon to the main channel. This site was characterized by muddy-sandy sediments, limited OM enrichment of sediments, abundant submerged vegetation (e.g., Ruppia), and a significantly higher hydrodynamics than at the other sites [47].
2.2. Field Surveysand Sample Treatment
The field surveys were carried out at sites C1, C2, and C3 on 6 July 2010 and 2 February 2011. At each site and date, water temperature, salinity, and dissolved oxygen (DO) were measured using portable probes (WTW LF 197 and WTWOxil 197, respectively). Subsequently, sediment samples for the determination of the water content (Wc) and chemical analysis (OM, chlorophyll-a, and phaeopigment content) were collected using a manual core (40 cm long, 5.5 cm diameter) gently pushed by hand into the sediments. Procedural details of sediment collection and chemical analysis are given in the companion paper by [30].
For the analyses of the meiofauna, six replicates were collected at each site by means of plexiglas corers (diameter: 3.6 cm) inserted 5 cm in the sediment. These samples were pre-filtered with magnesium chloride (MgCl2; 80 g L−1) to allow organisms to relax before fixation and facilitate subsequent taxonomic identification [48]. This treatment appears important because the “soft-bodied” taxa (e.g., Gastrotricha, Plathelminthes, and Nemertina) usually undergo the major morphological alterations after fixation and they can remain in good conditions with magnesium chloride treatment. The sediment samples were then fixed in a solution of pre-filtered seawater containing formalin buffered with sodium tetraborate Na2B4O7 to reach a pH of ca. 8.2 [49]. The amount of formalin to be added to the sample to obtain a final concentration of 4% was calculated based on the total volume of sediment and water present in the sample. A few drops of a Rose Bengal solution (0.5 g L−1) were added to the sample in order to facilitate the identification of organisms in the sorting phase [50].
2.3. Meiofaunal Analysis
The samples were rinsed with a gentle jet of fresh water through a 0.5 mm sieve to separate the macrofauna from the meiofauna [48]. They were then decanted, sieved 10 times through a 42 μm mesh and centrifuged three times with Ludox HS30 (specific density 1.18 g/cm3) [51]. The obtained animals were then transferred to a “Delfuss” Petri dish with a checkered bottom (200 squares, to make counting easier), sorted into their major taxa under a Leica G26 stereomicroscope, and counted.
All the values obtained was recalculated as abundance per 10 cm2. The richness (number of major taxa), Shannon’s diversity, Pielou’s evenness indices (log2) were calculated to describe the structure of the meiofaunal assemblage. The possible occurrence of anthropogenic impact on the meiofaunal community was also assessed by the total number of nematodes (Ne) and copepods (Co) computed in the Ne:Co ratio. This index was proposed by Raffaelli and Mason [52] for the pollution monitoring with meiofauna. The hypothesis was that the divergent auto-ecological characteristics of these two abundant and frequent meiofaunal components (the extreme tolerance of nematodes and the high sensitivity of copepods) might detect the occurrence of human stress in marine sediments.
2.4. Statistical Analysis
Both abiotic and biotic data were used for the data analysis. Water variables were temperature, salinity, and dissolved oxygen; sediment variables included water content, OM, chlorophyll-a, and phaeopigments. Biotic data consisted in the abundance of the meiofauna and were used to construct a taxa-by-site and period matrix. The environmental data variation was represented by means of box-plots for each variable and each site. The biotic parameters computed were the number of taxa (S, taxon richness), the number of individuals per taxa (A, abundance), and the Shannon (H’, diversity) and Pielou (J, evenness) indices, as well as Ne/Co ratio. These biotic variables were computed for the three sampling sites C1, C2, and C3, for each replicate and date.
As for multivariate analysis, the non-parametric permutational analysis of variance (two way-PERMANOVA), based on Bray-Curtis (dis)similarity measures [53] was carried out to test significant differences of the structure of community among sites (three levels: C1, C2, and C3), periods (two levels: July and February), and site × period interactions as fixed factors. The data were log(x+1) transformed before the analysis. The PERMANOVA, based on Euclidean distance, was also used to test the significant differences of all the biotic univariate measures (i.e., total meiofaunal abundance, number of taxa, Shannon-diversity, Pielou evenness, and Ne/Co ratio). A log (x+1) transformation of data was applied only for the total meiofaunal abundance. The significance was computed by permutation with 9999 replicates. The pairwise comparisons between all pairs of sites were computed as post-hoc test and the Bonferroni correction procedure was followed to account for multiple simultaneous correlations [54]. The principal component analysis (PCA), based on the correlation matrix, was used to explore the faunal variations within the lagoon and periods. The environmental variables were used to understand the key environmental variables accounting for the much % of variance affecting the meiofaunal distribution. The multivariate procedure non-metric multidimensional scaling (nMDS) was used to investigate the differences between the sites; the more informative environmental variables were added in the analysis to best explain the meiobenthos structure and they were superimposed in the graph [55]. The meiofaunal major taxa contributing most to (dis)similarities among the sites were identified using the similarity percentages (SIMPER) test.
3. Results
3.1. Environmental Variables
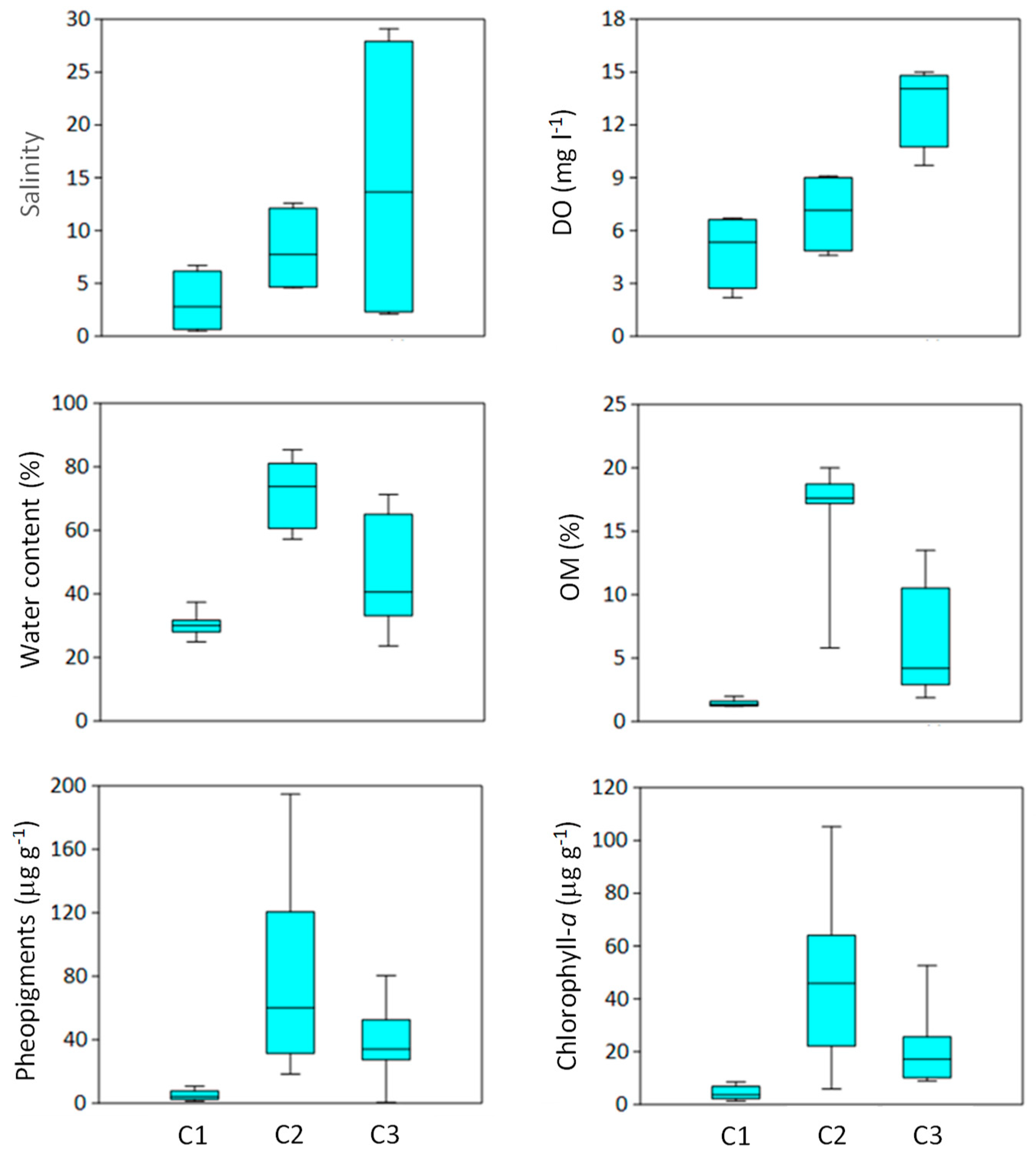
Among the water variables, both salinity and DO showed increasingly higher values from C1 to C3 (Figure 2), most variation found in salinity at C3 due to large differences between the summer (27 psu) and winter (3 psu) dates [30]. Differently from salinity, the median of DO was significantly higher at C3 than at both C1 and C2. Sediment variables had large within- and between-site variation. In particular, while C1 had low and homogeneous Wc, OM, chlorophyll-a (Chl-a), and phaeopigment values, C2 was most variable and had the highest peaks and outliers in OM, Chl-a, and phaeopigments among the three sites. C3 was in an intermediate position, with the medians of all sediment variables between C1 and C2 (Figure 2).
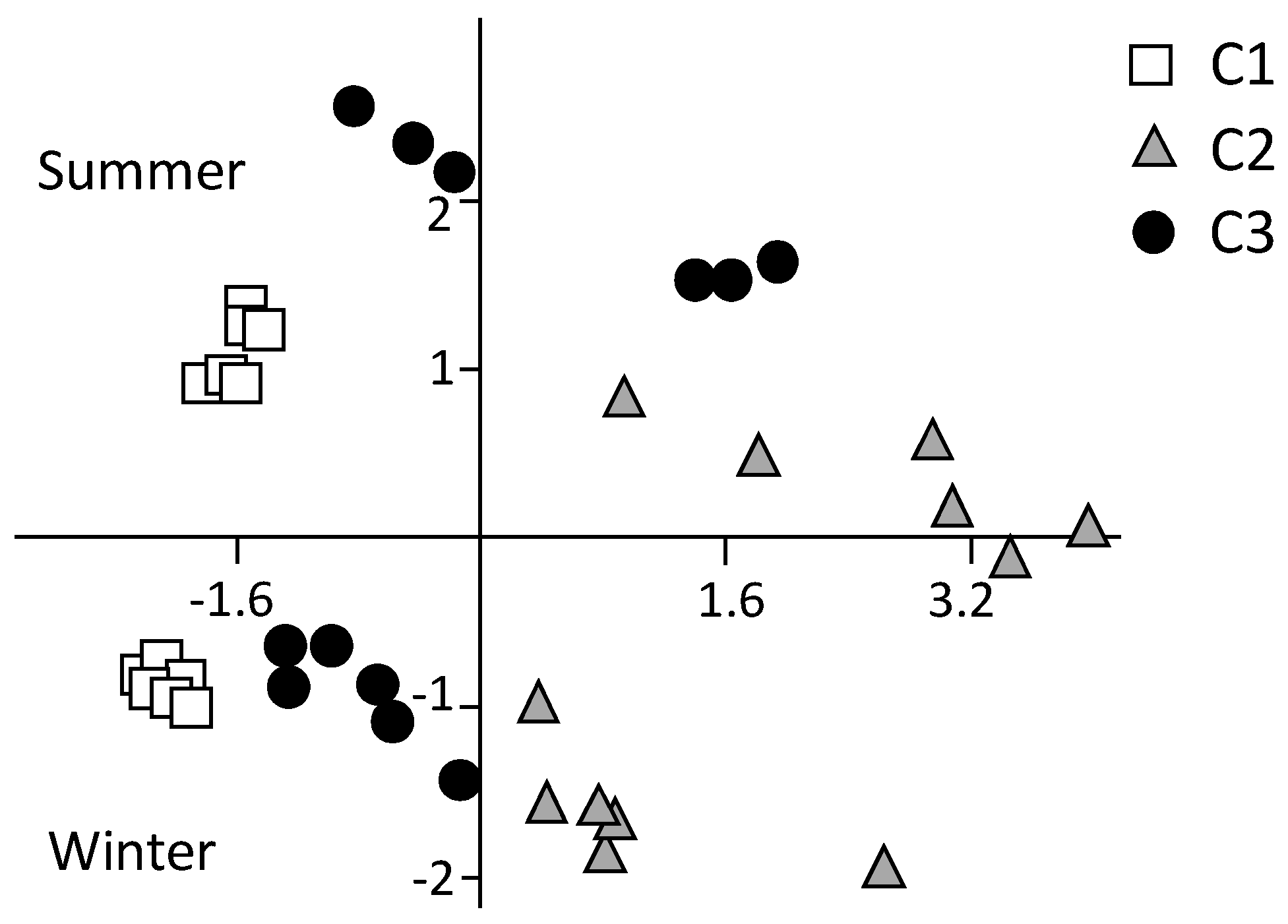
The PCA showed a clear separation of both sites and dates (Figure 3). C1 data-points were located on the left-had side of the ordination model (second and third quadrants), while those of C2 were located on the right-hand side (first and fourth quadrants). C3 was in intermediate position, yet clearly separated from C1 and C2. The two sampling dates were also clearly distinguishable for each site, summer and winter data-point being positioned on the upper and lower portion of the model, respectively. Among the environmental variables, sediment Wc, OM, Chl-a, and phaeopigments were mostly correlated with PC1, while water temperature and salinity showed the highest correlation with PC2 (Table 1). This indicates that sites were discriminated on the x-axis by the sediment variables mostly affected by the trophic features, while on the y-axis they were distributed in relation to the confinement gradient as indicated by the water variables. The variance explained by the model was 46.1% and 23.5% for PC1 and PC2, respectively.
3.2. Meiofauna
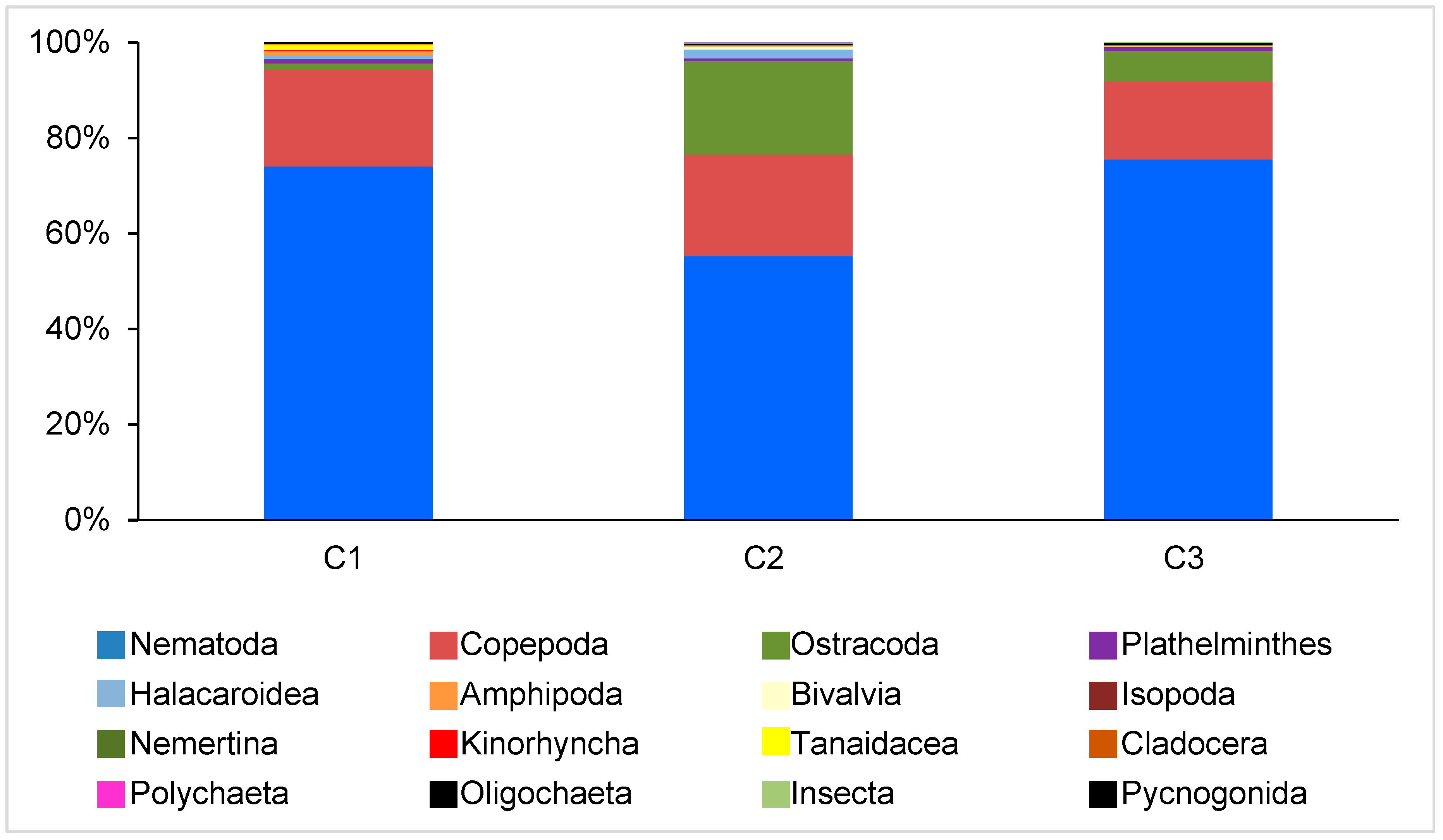
A total of 16 meiofaunal taxa were found: Plathelminthes, Nemertina, Nematoda, Kinorhyncha, Bivalvia, Polychaeta (adults and nectochaetes), Oligochaeta, Copepoda (adults and juveniles), Ostracoda, Amphipoda, Cladocera, Isopoda, Tanaidacea, Insecta, Halacaroidea, and Pycnogonida. The dominant taxa were generally nematodes (from 41% at C2 in winter 2011 to 86% at C3 in summer 2010), copepods (from 4% at C3 in summer 2010 to 33% C3 in winter 2011), and ostracods (from 0% at C2 in summer 2010 to 12% at C3 in winter 2011).
The PERMANOVA carried out on the structure of the meiofaunal assemblage indicated highly significant differences only among sites (p < 0.001), while no significant differences emerged among periods or site × period interactions (Table 2). In particular, pairwise comparisons highlighted significant differences between C3 and the other two sites (C1 and C2). The aforementioned variations were tested by means of the SIMPER analysis, which highlighted that five main groups, i.e., Nematoda, Copepoda, Ostracoda, Nauplii, and Halacaroidea, contributed up to the 95% of the cumulative similarity, with an individual contribution varying from 58% for Nematoda to 2.3% for Halacaroidea (Table 3; Figure 4). All the other taxa contributed to less than 2%.
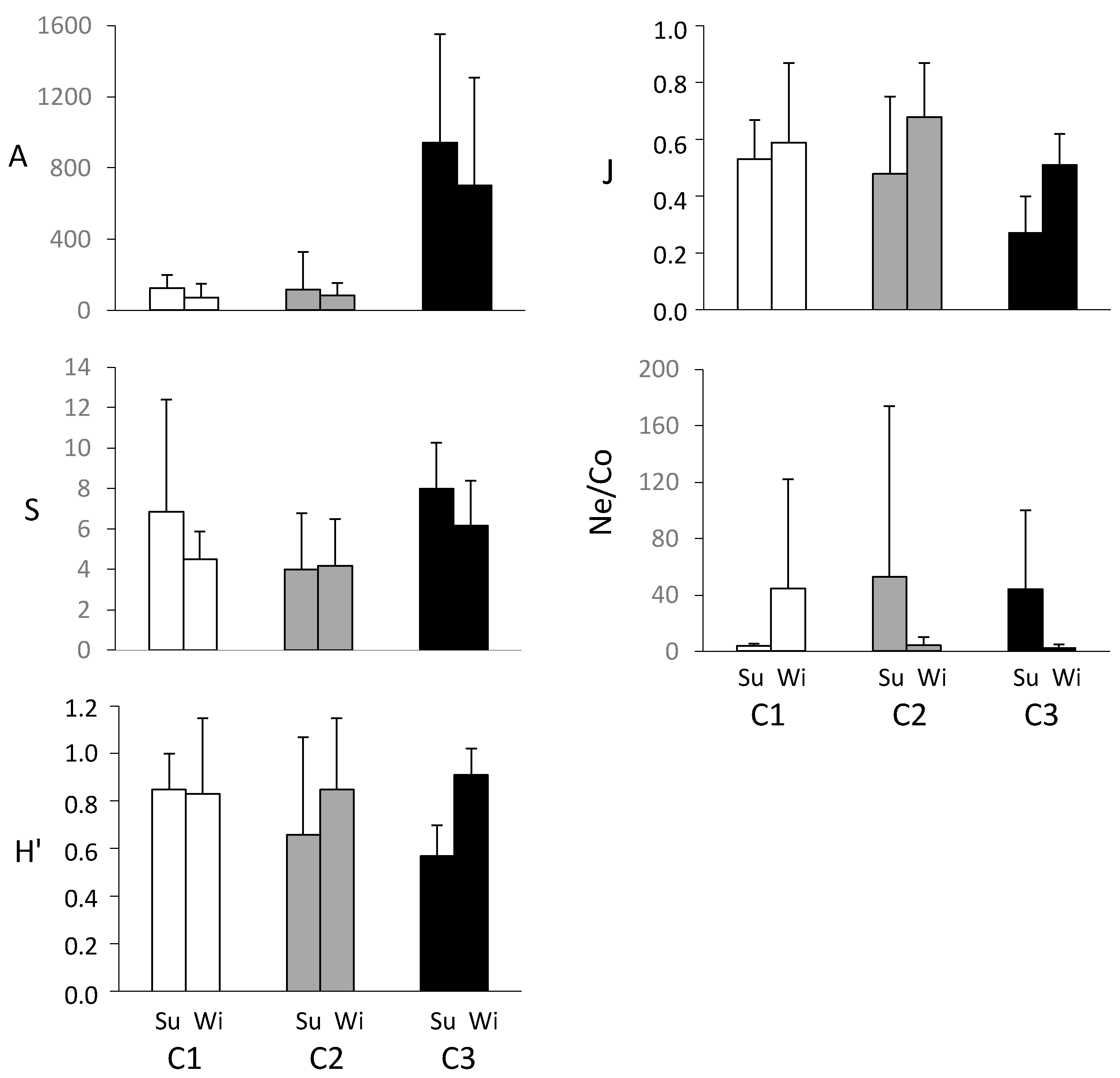
Variation in the meiofaunal abundance, number of taxa, diversity, evenness, and the Ne/Co ratio are shown in Figure 5. While differences between sites in abundance, evenness, and the Ne/Co ratio were high, those in taxon number and diversity were not. In particular, abundance was the highest at C3, which also showed the most marked standard error, and consequently the lowest evenness. The Ne/Co ratio was higher in summer, but always lower in winter both at C2 and C3, which coincided, with the drop in salinity. Finally, the most confined and organically enriched site C2 showed the most variation in the Ne/Co ratio up to various orders of magnitude, particularly in summer. The 2-way PERMANOVA conducted individually on each biotic measure showed significant differences among sites in the total abundance, taxon number, evenness and the Ne/Co ratio, while no difference was found for the diversity index (Table 4). The pairwise comparisons among the three sites are showed in Table 4 and highlighted higher and significantly differences especially of C3. PERMANOVA did not reveal significant differences of meiofaunal abundance, taxon richness, Shannon diversity, and Ne/Co ratio among the periods investigated, while some differences of evenness (p < 0.05) were found (Table 4).
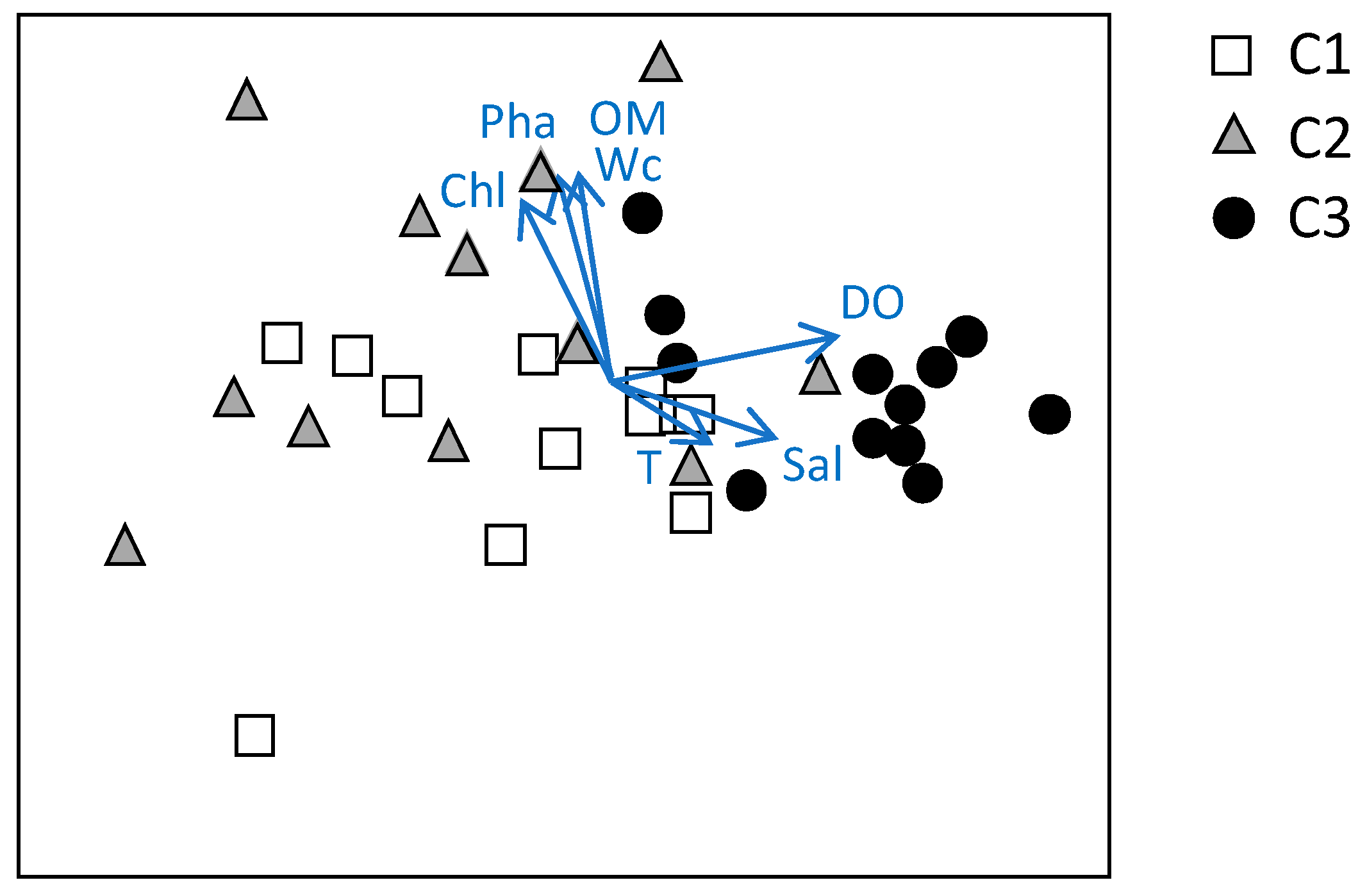
The nMDS on the structure of assemblage similarly highlighted three major groups corresponding to the three study sites, with a partial overlap between C1 and C2, and a major separation of C3 from both C1 and C2 (Figure 6). In this analysis, the environmental variables superimposed to the biotic data showed that salinity and DO were the main responsible factors for the separation of C3 from C1 and C2 (on the right-hand side) and that Wc, OM, Chl-a, and phaeopigments contributed mostly to the separation between C1 and C2 (Figure 6). These results demonstrated two main gradients influencing the spatiotemporal variation in the meiofaunal assemblages in the Cabras Lagoon. Temperature, on the contrary, did not make a relevant contribution to the ordination of the sites as indicated by its short segment.
4. Discussion
The present study provides the first information of the spatiotemporal variation in lagoonal meiofaunal assemblages in one of the richest Italian regions in number and extension of lagoons within the Tyrrhenian coast, i.e., the Sardinian Island. We demonstrated significant changes in meiofaunal diversity and community structure in relation to the main environmental gradients which, in the studied lagoon, are driven by salinity and dissolved oxygen concentration in water, sediment organic enrichment, and different degrees of confinement and saprobity. Our results highlight the importance of meiofaunal studies often neglected, by ecologists and policy makers, to further our understanding of the functioning of Mediterranean lagoons.
From an abiotic point of view, the heterogeneity of the Cabras Lagoon can be related to two main interlinked environmental and ecological gradients. The first gradient is due to the marine influence seaward, the second one is related to spatial differences in the trophic status and saprobity condition across the lagoon. In particular, the most confined site C2 differed from the other two investigated sites as being characterized by muddy, organically enriched sediments, and due to its lack of connection and exchange with both continental and marine waters, mostly influencing C1 and C3, respectively. Differently, the northern site C1 was most directly affected by the riverine input of freshwater and had sandy sediments with a very low phytopigment and OM content. This is consistent with the high energy and the low water residence time found at C1, which helps explaining the resuspension and export of fine sediment particles from this site to the central sector of the lagoon [56,57]. Finally, the seaward site C3 located in one of the three creeks connecting the lagoon to the Gulf of Oristano was in terms of sediment features intermediate between C1 and C2. It is worthwhile noting that in this sector of the lagoon man-made structures constructed in proximity of the inlet have caused modifications in the sedimentary regime of the lagoon and the water exchange with the adjacent Gulf [41,56]. Yet, the largest variation in salinity was found at C3 due to a marked drop in salinity in the whole lagoon in the winter of the present study, nullifying temporarily the salinity gradient and further highlighting the high seasonal and interannual environmental variability of the Cabras Lagoon [35,36].
Overall, these results show that the environmental (dis)similarities between the three study sites were constant in the two periods investigated, although summer and winter data-points were most clumped at C1 and more widespread, but still clearly separated one another, at C2 and C3. Thus, temporal variation did not alter significantly the location of the site-points in the multivariate model ordination. This highlighted that the most relevant source of variation in the study area was not due to differences between summer and winter dates, but to differences between sampling sites.
Regarding the meiofaunal richness, our data can be compared only with a few studies because the complete list of meiofaunal taxa is rarely reported. However, an overall high number of richness (i.e., 16 taxa) was recorded in the present study showing levels higher than those documented from both Mediterranean and European transitional water bodies [17,18,19,22,58,59]. Nematodes were the dominant taxon as frequently reported in lagoon systems worldwide [2,17,22,60,61]. This is likely related to their capacity to colonize the fine (suboxic or anoxic) sediments that generally characterize lagoons [19]. As reported in literature, the second most abundant group is represented by copepods [14,22,58,59]. Copepods are one of the most sensitive taxa to oxygen limitation and therefore they are confined to the oxic sediments [20,48], but they seem to take advantage of the high abundance of the microphytobenthos occurring in lagoon sediments and that are a primary food source for numerous copepod species [24].
The meiofaunal structure assemblage seemed to be very sensitive to the spatial environmental heterogeneity found in the Cabras Lagoon appearing a promising indicator of biotic changes in transitional water bodies. Indeed, the multivariate analysis (nMDS and PERMANOVA) applied on the assemblage structure clearly distinguished the three different sites in line with their environmental features with a partial overlap of the meiofaunal structure of C1 and C2, and a greater separation of C3 (Table 2, Figure 6). The latter site, characterized by “marine conditions”, had the highest abundance of all the meiofaunal taxa and in particular, it was distinguished by the greater abundances of Plathelminthes and Oligochaeta. These two taxa are often closely associated with each other and are among the primary components of transitional environment sediments [20,24]. Oligochaeta are regarded as taxa able to adapt to numerous environmental stress [18,48]. Plathelminthes are effective predators of many meiobenthic organisms such as copepods that could explain their higher abundance at C3 where the higher meiofaunal densities and copepods was found [14,62].
Instead, the high degree of confinement and consequent trophic load existing at site C2 was marked by a higher presence of Halacaroidea and Ostracoda. Ostracoda are generally recognized as sensitive taxon to environmental perturbations, but adaptive behaviors to numerous natural and anthropogenic environmental (e.g., organic load and trace element contamination) changes have been documented in several species [22,24,63]. In that site, there was also the only record of Pycnogonida that is generally recognized as a marine taxon. Pycnogonida have a few representatives in the meiofauna, but the Anoplodactylus genus has some species that ranges in the meiofaunal body size and are also tolerant to salinity variations until values comparable to those found in C2 (11 PSU) [64].
In C1, the site with the lowest salinity, Tanaidacea was one of the discriminating taxon. Noteworthy is that although many euryhaline species from Tanaidacea are found in transitional habitats, most occur only temporarily in these environments, appearing unable to form stable populations there [65]. Furthermore, Ateş et al. [66] reported that some species are particularly related to coarse grain size and low content of organic matter that were the conditions that distinguish C1.
The environmental variables that appeared to mainly affect the meiofaunal assemblage of the study area were salinity and DO that were the main responsible parameters for the separation of C3 and the other two sites. Salinity gradient is one of the primary factors that influence meiofauna in transitional environments [8,18] along with the oxygen availability that seems to influence all the meiofaunal taxa and not only the oxygen sensitive ones such as copepods (see above references). The quantity and quality of the organic matter (OM, Chl-a, and phaeopigments) and Wc, which is an indirect indication of the grain size of the substrates, were also important for the meiofaunal distribution as suggested by many authors [8,17,22] and contributed mostly to the separation between the other two sites (i.e., C1 and C2). Instead, temperature did not show a relevant contribution to the ordination of the meiofauna. This issue as well as the lack of significant differences in the comparison of the meiofaunal assemblage structure suggest that meiofauna was more affected by spatial than temporal variations. This pattern resembled the distribution patterns of the macrozoobenthos observed at the same study sites where spatial differences were greater than significant seasonal changes [30].
When the spatiotemporal variation in the univariate measures were statistically studied, the greatest differences were observed between the three sites further supporting the results revealed by the structure of the meiofaunal assemblage (Table 4). In particular, PERMANOVA showed significant differences of the total meiofaunal abundance, number of taxa, evenness and the Ne/Co ratio. Among them, the Ne/Co ratio showed a very temporal variable trend (i.e., it was higher in C1 in the winter and lower in C2 and C3 in the summer) which likely is why PERMANOVA did not reveal significant differences between periods. However, the variation in the Ne/Co ratio did not highlight the presence of anthropogenic stress being the values lower than the thresholds reported by Raffaelli and Mason [52] for stressful conditions. Instead, the highest evenness at C1 was likely related to the coarser grain size of the sediments that generally host a more diversified meiofaunal assemblage [67,68].
5. Conclusions
The present study is one of the few investigations on the meiofaunal community structure, composition, and diversity conducted in transitional waters (TWs) in the Western Mediterranean Sea. The high environmental variability and habitat heterogeneity of the Cabras Lagoon, the largest TW in Sardinia, was reflected in significant differences in meiofauna among the study sites. Spatial differences in several faunal parameters (i.e., community structure, richness, Pielou-evenness, and Ne/co ratio) were stronger than temporal variation, suggesting that meiofaunal organisms are good indicators of the physical-chemical variation in TWs. Furthermore, the Cabras Lagoon showed high values of meiofaunal species richness further supporting the idea that TWs may be biodiversity hotspots and meiofauna is an important biotic component to understand their functioning.
Author Contributions
Conceptualization, F.S., M.F.G. and P.M.; methodology, F.S., M.F.G. and P.M.; formal analysis, F.S., M.F.G. and P.M.; data curation, F.S., M.F.G. and P.M.; writing-original draft preparation, F.S., M.F.G. and P.M.; writing-review and editing, F.S., M.F.G. and P.M.; visualization, F.S., M.F.G. and P.M.
Funding
We would like to thank Marcello Giorgi, University of Rome “Tor Vergata”, for technical support in data analysis procedures.
Acknowledgments
We gratefully acknowledge the two reviewers whose comments greatly contributed to an improved version of the original manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cardone, F.; Corriero, G.; Fianchini, A.; Gravina, M.F.; Nonnis Marzano, C. Biodiversity of transitional waters: Species composition and comparative analysis of hard bottom communities from south-eastern Italian coast. J. Mar. Biol. Assoc. UK 2014, 94, 25–34. [Google Scholar] [CrossRef]
- Kandratavicius, N.; Muniz, P.; Venturini, N.; Giménez, L. Meiobenthic communities in permanently open estuaries and open/closed coastal lagoons of Uruguay (Atlantic coast of South America). Estuar. Coast. Shelf Sci. 2015, 163, 44–53. [Google Scholar] [CrossRef]
- Barnes, R.S.K. Coastal Lagoons: The Natural History of a Neglected Habitat; Cambridge University Press: Cambridge, UK, 1980; pp. 1–180. [Google Scholar]
- Pérez-Ruzafa, A.; Marcos, C.; Pérez-Ruzafa, I.M. Mediterranean coastal lagoons in an ecosystem and aquatic resources management context. Phys. Chem. Earth 2011, 36, 160–166. [Google Scholar]
- Magni, P.; Tagliapietra, D.; Lardicci, C.; Balthis, L.; Castelli, A.; Como, S.; Frangipane, G.; Giordani, G.; Hyland, J.; Maltagliati, F.; et al. Animal-sediment relationships: Evaluating the ‘Pearson-Rosenberg paradigm’ in Mediterranean coastal lagoons. Mar. Pollut. Bull. 2009, 58, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Armynot du Châtelet, E.; Bout-Roumazeilles, V.; Coccioni, R.; Frontalini, F.; Francescangeli, F.; Margaritelli, G.; Rettori, R.; Spagnoli, F.; Semprucci, F.; Trentesaux, A.; et al. Environmental control on a land-sea transitional setting–Integrated microfaunal, sedimentological, and geochemical approaches. Environ. Earth Sci. 2016, 75, 123. [Google Scholar] [CrossRef]
- Bouchet, V.M.P.; Goberville, E.; Frontalini, F. Benthic foraminifera to assess Ecological Quality Status in Italian transitional waters. Ecol. Indic. 2018, 84, 130–139. [Google Scholar] [CrossRef]
- Gambi, C.; Totti, C.; Manini, E. Impact of organic loads and environmental gradients on microphytobenthos and meiofaunal distribution in a coastal lagoon. Chem. Ecol. 2003, 19, 207–223. [Google Scholar] [CrossRef]
- Moens, T.; Braeckman, U.; Derycke, S.; Fonseca, G.; Gallucci, F.; Ingels, J.; Leduc, D.; Vanaverbeke, J.; Van Colen, C.; Vanreusel, A.; et al. Ecology of free-living marine nematodes. In Handbook of Zoology: Gastrotricha, Cycloneuralia and Gnathifera Nematoda; Schmidt-Rhaesa, A., Ed.; deGruyter: Berlin, Germany, 2013; Volume 2, pp. 109–152. [Google Scholar]
- Boufahja, F.; Semprucci, F.; Beyrem, H. An experimental protocol to select nematode species from an entire community using progressive sedimentary enrichment. Ecol. Indic. 2016, 60, 292–309. [Google Scholar] [CrossRef]
- Baldrighi, E.; Semprucci, F.; Franzo, A.; Cvitkovic, I.; Bogner, D.; Despalatovic, M.; Berto, D.; MalgorzataFormalewicz, M.; Scarpato, A.; Frapiccini, E.; et al. Meiofaunal communities in four Adriatic ports: Baseline data for risk assessment in ballast water management. Mar. Pollut. Bull. 2018. [Google Scholar] [CrossRef]
- Yang, X.; Lin, C.; Song, X.; Xu, M.; Yang, H. Effects of artificial reefs on the meiofaunal community and benthic environment—A case study in Bohai Sea, China. Mar. Pollut. Bull. 2019, 140, 179–187. [Google Scholar] [CrossRef]
- Semprucci, F.; Balsamo, M. Free-living Marine Nematodes as Bioindicators: Past, Present and Future Perspectives. Trends Environ. Sci. 2014, 6, 17–36. [Google Scholar]
- Colangelo, M.A.; Ceccherelli, V.U. Meiofaunal recolonization of azoicsediment in a Po Delta lagoon (Sacca di Goro). Ital. J. Zool. 1994, 61, 335–342. [Google Scholar]
- Villano, N.; Warwick, R.M. Meiobenthic communities associated with the seasonal cycle of growth and decay of Ulva rigidaArardh in the Palude Della Rosa, Lagoon of Venice. Estuar. Coast. Shelf. Sci. 1995, 4, 181–194. [Google Scholar] [CrossRef]
- Guerrini, A.; Colangelo, M.A.; Ceccherelli, V.U. Recolonization patterns of meiobenthic communities in brackish vegetated and unvegetated habitats after induced hypoxia/anoxia. Hydrobiologia 1998, 375–376, 73–87. [Google Scholar] [CrossRef]
- Pusceddu, A.; Gambi, C.; Manini, E.; Danovaro, R. Trophic state, ecosystem efficiency and biodiversity of transitional aquatic ecosystems: Analysis of environmental quality based on different benthic indicators. Chem. Ecol. 2007, 23, 1–11. [Google Scholar] [CrossRef]
- Cibic, T.; Franzo, A.; Celussi, M.; Fabbro, C.; Del Negro, P. Benthic ecosystem functioning in hydrocarbon and heavy-metal contaminated sediments of an Adriatic lagoon. Mar. Ecol. Prog. Ser. 2012, 458, 69–87. [Google Scholar] [CrossRef] [Green Version]
- Semprucci, F.; Balsamo, M.; Sandulli, R. Assessment of the Ecological quality (EcoQ) of the Venice lagoon using the structure and biodiversity of the meiofaunal assemblages. Ecol. Indic. 2016, 67, 451–457. [Google Scholar] [CrossRef]
- Semprucci, F.; Facca, C.; Ferrigno, F.; Balsamo, M.; Sfriso, A.; Sandulli, R. Biotic and abiotic factors affecting seasonal and spatial distribution of meiofauna and macrophytobenthos in transitional coastal waters. Estuar. Coast. Shelf Sci. 2019, 219, 328–340. [Google Scholar] [CrossRef]
- Fabbrocini, A.; Guarino, A.; Scirocco, T.; Franchi, M.; D’Adamo, R. Integrated biomonitoring assessment of the Lesina Lagoon (Southern Adriatic Coast, Italy): Preliminary results. Chem. Ecol. 2005, 21, 479–489. [Google Scholar] [CrossRef]
- Frontalini, F.; Semprucci, F.; Armynot du Châtelet, E.; Francescangeli, F.; Margaritelli, G.; Rettori, R.; Spagnoli, F.; Balsamo, M.; Coccioni, R. Biodiversity trends of the meiofauna and foraminifera assemblages of Lake Varano (southern Italy). Proc. Biol. Soc. Wash. 2014, 127, 7–22. [Google Scholar] [CrossRef]
- Semprucci, F.; Balsamo, M.; Frontalini, F. The nematode assemblage of a coastal lagoon (Lake Varano, Southern Italy): Ecology and biodiversity patterns. Sci. Mar. 2014, 78, 579–588. [Google Scholar] [CrossRef]
- Mirto, S.; La Rosa, T.; Morcciaro, G.; Costa, K.; Sara, G.; Mazzola, A. Meiofauna and benthic microbial biomass in a semi-enclosed Mediterranean marine system (Stagnone of Marsala, Italy). Chem. Ecol. 2004, 20, S387–S396. [Google Scholar] [CrossRef]
- Manini, E.; Fiordelmondo, C.; Gambi, M.C.; Pusceddu, A.; Danovaro, R. Benthic microbial loop functioning in coastal lagoons: A comparative approach. Oceanol. Acta 2003, 26, 27–38. [Google Scholar] [CrossRef]
- Barnes, N.; Bamber, R.; Moncrieff, C.; Sheader, M.; Ferrero, T. Meiofauna in closed coastal saline lagoons in the United Kingdom: Structure and biodiversity of nematode assemblage. Estuar. Coast. Shelf Sci. 2008, 79, 328–340. [Google Scholar] [CrossRef]
- Abbiati, M.; Mistri, M.; Bartoli, M.; Ceccherelli, V.U.; Colangelo, M.A.; Ferrari, C.R.; Giordani, G.; Munari, C.; Nizzoli, D.; Ponti, M.; et al. Trade-off betweenconservation and exploitation of the transitional water ecosystems of the northernAdriatic Sea. Chem. Ecol. 2010, 26, 105–119. [Google Scholar] [CrossRef]
- Magni, P.; Micheletti, S.; Casu, D.; Floris, A.; Giordani, G.; Petrov, A.; De Falco, G.; Castelli, A. Relationships between chemical characteristics of sediments and macrofaunal communities in the Cabras lagoon (western Mediterranean, Italy). Hydrobiologia 2005, 550, 109–115. [Google Scholar] [CrossRef]
- Magni, P.; Rajagopal, S.; van der Velde, G.; Fenzi, G.; Kassenberg, J.; Vizzini, S.; Mazzola, A.; Giordani, G. Sediment features, macrozoobenthic assemblages and trophic relationships (δ13C and δ15N analysis) following a dystrophic event with anoxia and sulphide development in the Santa Giusta lagoon (western Sardinia, Italy). Mar. Pollut. Bull. 2008, 57, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Foti, A.; Fenzi, G.; Di Pippo, F.; Gravina, M.F.; Magni, P. Testing the saprobity hypothesis in a Mediterranean lagoon: Effects of confinement and organic enrichment on benthic communities. Mar. Environ. Res. 2014, 99, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Ferrarin, C.; Umgiesser, G. Hydrodynamic modeling of a coastal lagoon: The Cabras lagoon in Sardinia, Italy. Ecol. Model. 2005, 188, 340–357. [Google Scholar] [CrossRef]
- Molinaroli, E.; Guerzoni, S.; De Falco, G.; Sarretta, A.; Cucco, A.; Como, S.; Simeone, S.; Perilli, A.; Magni, P. Relationships between hydrodynamic parameters and grain size in two contrasting transitional environments: The lagoons of Venice and Cabras, Italy. Sediment. Geol. 2009, 219, 196–207. [Google Scholar] [CrossRef]
- Bartoli, M.; Longhi, D.; Nizzoli, D.; Como, S.; Magni, P.; Viaroli, P. Short term effects of hypoxia and bioturbation on solute fluxes, denitrification and buffering capacity in a shallow dystrophic pond. J. Exp. Mar. Biol. Ecol. 2009, 381, 105–113. [Google Scholar] [CrossRef]
- Specchiulli, A.; Cilenti, L.; D’Adamo, R.; Fabbrocini, A.; Guo, W.; Huang, L.; Lugliè, A.; Padedda, B.M.; Scirocco, T.; Magni, P. Dissolved organic matter dynamics in Mediterranean lagoons: The relationship between DOC and CDOM. Mar. Chem. 2018, 202, 37–48. [Google Scholar] [CrossRef]
- Pulina, S.; Padedda, B.M.; Satta, C.T.; Sechi, N.; Lugliè, A. Long-term phytoplankton dynamics in a Mediterranean Eutrophic lagoon (Cabras Lagoon, Italy). Plant Biosyst. 2011, 146, 259–272. [Google Scholar] [CrossRef]
- Pulina, S.; Padedda, B.M.; Sechi, N.; Lugliè, A. The dominance of cyanobacteria in Mediterranean hypereutrophic lagoons: A case study of Cabras lagoon (Sardinia, Italy). Sci. Mar. 2012, 75, 111–120. [Google Scholar] [CrossRef]
- Magni, P.; Micheletti, S.; Casu, D.; Floris, A.; De Falco, G.; Castelli, A. Macrofaunal community structure and distribution in a muddy coastal lagoon. Chem. Ecol. 2004, 20, S397–S407. [Google Scholar] [CrossRef]
- Satta, C.T.; Anglès, S.; Garcés, E.; Sechi, N.; Pulina, S.; Padedda, B.M.; Stacca, D.; Lugliè, A. Dinoflagellate cyst assemblages in surface sediments from three shallow Mediterranean lagoons (Sardinia, North Western Mediterranean Sea). Estuaries Coasts 2014, 37, 646–663. [Google Scholar] [CrossRef]
- Como, S.; Magni, P.; Van Der Velde, G.; Blok, F.S.; Van De Steeg, M.F.M. Spatial variations in δ13C and δ15N values of primary consumers in a coastal lagoon. Estuar. Coast. Shelf Sci. 2012, 115, 300–308. [Google Scholar] [CrossRef]
- Cucco, A.; Sinerchia, M.; Le Francois, C.; Magni, P.; Ghezzo, M.; Umgiesser, G.; Perilli, A.; Domenici, P. Coupled empirical and numerical model of fish response to environmental changes. Ecol. Model. 2012, 237–238, 132–141. [Google Scholar] [CrossRef]
- Como, S.; Magni, P.; Casu, D.; Floris, A.; Giordani, G.; Natale, S.; Fenzi, G.A.; Signa, G.; De Falco, G. Sediment characteristics and macrofauna distribution along a human-modified inlet in the Gulf of Oristano (Sardinia, Italy). Mar. Pollut. Bull. 2007, 54, 733–744. [Google Scholar] [CrossRef]
- Como, S.; Magni, P. Temporal changes of a macrobenthic assemblage in harsh lagoon sediments. Estuar. Cost. Shelf Sci. 2009, 83, 638–646. [Google Scholar] [CrossRef]
- Sandulli, R.; De Leonardis, C.; Vanaverbeke, J. Meiobenthic communities in the shallow subtidal of three Italian Marine Protected Areas. Ital. J. Zool. 2010, 77, 186–196. [Google Scholar] [CrossRef] [Green Version]
- Curini-Galletti, M.; Artois, T.; Delogu, V.; De Smet, W.H.; Fontaneto, D.; Jondelius, U.; Leasi, F.; Martinez, A.; Meyer-Wachsmuth, I.; Nilsson, K.S.; et al. Patterns of Diversity in Soft-Bodied Meiofauna: Dispersal Ability and Body Size Matter. PLOS ONE 2012, 7, e33801. [Google Scholar] [CrossRef] [PubMed]
- Di Pippo, F.; Ellwood, N.T.W.; Gismondi, A.; Bruno, L.; Rossi, F.; Magni, P.; De Philippis, P. Characterization of biofilm-forming cyanobacteria for exopolysaccharide production and potential biotechnological applications. J. Appl. Phycol. 2013, 25, 1697–1708. [Google Scholar] [CrossRef]
- Di Pippo, F.; Magni, P.; Congestri, R. Microphytobenthic biomass, diversity and exopolymeric substances in a shallow dystrophic coastal lagoon. J. Mar. Microbiol. 2018, 2, 6–12. [Google Scholar]
- Magni, P.; Como, S.; Cucco, A.; De Falco, G.; Domenici, P.; Ghezzo, M.; Lefrançois, C.; Simeone, S.; Perilli, A. A Multidisciplinary and Ecosystemic Approach in the Oristano Lagoon-Gulf System (Sardinia, Italy) as a Tool in Management Plans. Transit. Waters Bull. 2008, 2, 41–62. [Google Scholar]
- Danovaro, R.; Gambi, M.C.; Mirto, S.; Sandulli, R.; Ceccherelli, V.U. Meiofauna. Biol. Mar. Mediterr. 2004, 11, 55–97. [Google Scholar]
- Higgins, R.P.; Thiel, H. Introduction to the Study of Meiofauna; Smithsonian Institution Press: Washington, DC, USA, 1988; pp. 1–488. [Google Scholar]
- Semprucci, F.; Sbrocca, C.; Rocchi, M.; Balsamo, M. Temporal changes of the meiofaunal assemblage as a tool for the assessment of the ecological quality status. J. Mar. Biol. Assoc. UK 2015, 95, 247–254. [Google Scholar] [CrossRef]
- Pfannkuche, O.; Thiel, H. Sample processing. In Introduction to the Study of Meiofauna; Higgins, R.P., Thiel, H., Eds.; Smithsonian Institute: Washington, DC, USA, 1988; pp. 134–145. [Google Scholar]
- Raffaelli, D.G.; Mason, D.F. Pollution monitoring with meiofauna, using the ratio of nematodes to copepods. Mar. Pollut. Bull. 1981, 12, 158–163. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Aust. Ecol. 2001, 26, 32–46. [Google Scholar]
- Rice, W.R. Analyzing tables of statistical tests. Evolution 1989, 43, 223–225. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. Changes in Marine Communities: An Approach to Statistical Analysis and Interpretation; Plymouth Marine Laboratory: Plymouth, UK, 2001. [Google Scholar]
- De Falco, G.; Magni, P.; Teräsvuori, L.; Matteucci, G. Sediment grain size and organic carbon distribution in the Cabras lagoon (Sardinia, west Mediterranean). Chem. Ecol. 2004, 20, S367–S377. [Google Scholar] [CrossRef]
- Magni, P.; De Falco, G.; Como, S.; Casu, D.; Floris, A.; Petrov, A.N.; Castelli, A.; Perilli, A. Distribution and ecological relevance of fine sediments in organic-enriched lagoons: The case study of the Cabras lagoon (Sardinia, Italy). Mar. Pollut. Bull. 2008, 56, 549–564. [Google Scholar] [CrossRef] [PubMed]
- Smol, K.A.; Willems, J.C.; Govaere, R.; Sandee, A.J.J. Composition, distributionand biomass of meiobenthos in the Oosterschelde estuary (SW Netherlands). Hydrobiologia 1994, 282–283, 197–217. [Google Scholar] [CrossRef]
- Jouili, S.; Essid, N.; Semprucci, F.; Boufahja, F.; Nasri, A.; Beyrem, H. Environmental quality assessment of El Bibane lagoon (Tunisia) using taxonomical and functional diversity of meiofauna and nematodes. J. Mar. Biol. Assoc. UK 2017, 97, 1593–1603. [Google Scholar] [CrossRef]
- McArthur, V.E.; Koutsoubas, D.; Lampadariou, N.; Dounas, C. The meiofaunal community structure of a Mediterranean lagoon (Gialova lagoon, Ionian Sea). Helgol. Mar. Res. 2000, 54, 7–17. [Google Scholar] [CrossRef] [Green Version]
- Alves, A.S.; Adão, H.; Patrício, J.; Magalhães Neto, J.; Costa, M.J.; Marques, J.C. Spatial distribution of subtidal meiobenthos along estuarine gradients in two southern European estuaries (Portugal). J. Mar. Biol. Assoc. UK 2009, 89, 1529–1540. [Google Scholar] [CrossRef]
- Watzin, M.C. Interactions among temporary and permanent meiofauna: Observations on the feeding and behaviour of selected taxa. Biol. Bull. 1985, 169, 397–416. [Google Scholar] [CrossRef]
- Vandekerkhove, J.; Martens, K.; Rossetti, G.; Mesquita-Joanes, F.; Namiotko, T. Extreme tolerance to environmental stress of sexual and parthenogenetic resting eggs of Eucypris virens (Crustacea, Ostracoda). Freshw. Biol. 2013, 58, 237–247. [Google Scholar] [CrossRef]
- Alvarez, F.; Ojeda, M. First record of a sea spider (Pycnogonida) from an anchialine habitat. Lat. Am. J. Aquat. Res. 2018, 46, 219–224. [Google Scholar] [CrossRef]
- Jaume, D.; Boxshall, G.A. Global diversity of cumaceans & tanaidaceans (Crustacea: Cumacea & Tanaidacea) in freshwater. Hydrobiologia 2008, 595, 225–230. [Google Scholar]
- Ateş, A.S.; Katağan, T.; Sezgin, M.; Acar, S. The Response of Apseudopsis latreillii (Milne-Edwards, 1828) (Crustacea, Tanaidacea) to Environmental Variables in the Dardanelles. Turk. J. Fish. Aquat. Sci. 2014, 14, 113–124. [Google Scholar] [CrossRef]
- Vanaverbeke, J.; Gheskiere, T.; Steyaert, M.; Vincx, M. Nematode assemblages from subtidal sandbanks in theSouthern Bight of the North Sea: Effect of smallsedimentological differences. J. Sea Res. 2002, 48, 197–207. [Google Scholar] [CrossRef]
- Semprucci, F.; Balsamo, M.; Appolloni, L.; Sandulli, R. Assessment of ecological quality status along the Apulian coasts (Eastern Mediterranean Sea) based on meiobenthic and nematode assemblages. Mar. Biodivers. 2018, 48, 105–115. [Google Scholar] [CrossRef]
Figure 1.
Location of the study area (Cabras lagoon, western Sardinia, Italy) and sampling sites (C1, C2, and C3). Image source: Google Earth.
Figure 1.
Location of the study area (Cabras lagoon, western Sardinia, Italy) and sampling sites (C1, C2, and C3). Image source: Google Earth.

Figure 2.
Mean values (n = 4, ±SE) of water salinity and dissolved oxygen (DO) concentration, and mean values (n = 12, ±SE) of sediment water (Wc), phaeopigment (Pha), organic matter (OM), and chlorophyll-a (Chl-a) contents at the sampling sites C1, C2, and C3 during the study period.
Figure 2.
Mean values (n = 4, ±SE) of water salinity and dissolved oxygen (DO) concentration, and mean values (n = 12, ±SE) of sediment water (Wc), phaeopigment (Pha), organic matter (OM), and chlorophyll-a (Chl-a) contents at the sampling sites C1, C2, and C3 during the study period.

Figure 3.
Principal component analysis (PCA) on environmental variables.

Figure 4.
Meiofauna community structure in the sampling sites C1, C2, and C3.

Figure 5.
Mean values (n = 6, ±SE standard error) of meiofaunal community synthetic measures at the sampling sites C1, C2, and C3 in summer (Su) and winter (Wi).
Figure 5.
Mean values (n = 6, ±SE standard error) of meiofaunal community synthetic measures at the sampling sites C1, C2, and C3 in summer (Su) and winter (Wi).

Figure 6.
MDS on meiofaunal abundance (stress = 0.09) with superimposed environmental.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Component correlation coefficients in the PCA of the environmental variables and the first two components. 69.6% of variance explained by Principal Components 1 (46.1%) and 2 (23.5%). In bold, the highest loadings.
Table 1.
Component correlation coefficients in the PCA of the environmental variables and the first two components. 69.6% of variance explained by Principal Components 1 (46.1%) and 2 (23.5%). In bold, the highest loadings.
| Variable | PC 1 | PC 2 |
|---|---|---|
| Temperature | 0.17 | 0.71 |
| Salinity | 0.24 | 0.59 |
| Dissolved oxygen | 0.004 | −0.12 |
| Water content | 0.49 | −0.27 |
| OM | 0.50 | −0.22 |
| Chlorophyll-a | 0.47 | 0.06 |
| Phaeopigments | 0.45 | −0.12 |
Table 2.
Results of two-way PERMANOVA (Bray Curtis (dis)similarity based) testing the differences among sites, periods and their interactions for the meiofaunal assemblage structure. Df = degrees of freedom. Significant p values are marked, ** for p < 0.001.
Table 2.
Results of two-way PERMANOVA (Bray Curtis (dis)similarity based) testing the differences among sites, periods and their interactions for the meiofaunal assemblage structure. Df = degrees of freedom. Significant p values are marked, ** for p < 0.001.
| Factors | Total Sum of Squares | Df | Mean Square | Pseudo-F | p | Pairwise |
|---|---|---|---|---|---|---|
| Sites | 2.72 | 2 | 1.36 | 6.36 | 0.0001 ** | C1 vs. C3 p = 0.0003 **; C2 vs. C3 p = 0.0006 ** |
| Periods | 0.20 | 1 | 0.20 | 0.94 | 0.44 | |
| Sites × Periods | 0.66 | 2 | 0.33 | 1.55 | 0.12 | |
| Residual | 6.41 | 30 | 0.21 | |||
| Total | 9.99 | 35 |
Table 3.
Results of the Similarity Percentage (SIMPER) analysis showing the average dissimilarity (Av. Dissim.), % contribution of each taxon (Cont. %), % cumulative for the pair comparisons (Cum. %), and average abundances of each sites (Av. ab.). The overall average dissimilarity is 70.9, 76.0, and 82.6 for C1 vs. C2, C1 vs. C3, and C2 vs. C3, respectively.
Table 3.
Results of the Similarity Percentage (SIMPER) analysis showing the average dissimilarity (Av. Dissim.), % contribution of each taxon (Cont. %), % cumulative for the pair comparisons (Cum. %), and average abundances of each sites (Av. ab.). The overall average dissimilarity is 70.9, 76.0, and 82.6 for C1 vs. C2, C1 vs. C3, and C2 vs. C3, respectively.
| C1 vs. C2 | Av. Dissim. | Cont. % | Cum. % | Av.ab. C1 | Av.ab. C2 | C1 vs. C3 | Av. Dissim. | Cont. % | Cum. % | Av.ab. C1 | Av.ab. C3 | C2 vs. C3 | Av. Dissim. | Cont. % | Cum. % | Av.ab. C2 | Av.ab. C3 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Nematoda | 41.2 | 58.1 | 58.1 | 73.1 | 54.8 | Nematoda | 52.8 | 69.4 | 69.4 | 73.1 | 619 | Nematoda | 58.0 | 70.2 | 70.2 | 54.8 | 619 |
| Copepoda | 13.4 | 18.9 | 77.0 | 17.5 | 20.3 | Copepoda | 12.5 | 16.4 | 85.8 | 17.5 | 89.8 | Copepoda | 13.0 | 15.8 | 85.9 | 20.3 | 89.8 |
| Ostracoda | 9.4 | 13.2 | 90.3 | 1.3 | 19.4 | Ostracoda | 4.9 | 6.5 | 92.3 | 1.3 | 53.4 | Ostracoda | 5.6 | 6.8 | 92.8 | 19.4 | 53.4 |
| Nauplii | 2.4 | 3.4 | 93.6 | 2.5 | 0.9 | Nauplii | 3.7 | 4.8 | 97.1 | 2.5 | 43.3 | Nauplii | 3.8 | 4.6 | 97.3 | 0.9 | 43.3 |
| Halacaroidea | 1.7 | 2.3 | 96.0 | 0.8 | 1.8 | Plathelminthes | 0.9 | 1.2 | 98.3 | 1.0 | 6.3 | Plathelminthes | 0.9 | 1.0 | 98.4 | 0.6 | 6.3 |
| Tanaidacea | 0.8 | 1.1 | 97.1 | 1.2 | 0.0 | Oligochaeta | 0.3 | 0.4 | 98.7 | 0.3 | 4.3 | Halacaroidea | 0.3 | 0.4 | 98.8 | 1.8 | 0.6 |
| Plathelminthes | 0.5 | 0.8 | 97.9 | 1.0 | 0.6 | Tanaidacea | 0.2 | 0.3 | 99.0 | 1.2 | 0.1 | Oligochaeta | 0.3 | 0.4 | 99.2 | 0.3 | 4.3 |
| Amphipoda | 0.3 | 0.5 | 98.3 | 0.7 | 0.0 | Amphipoda | 0.2 | 0.2 | 99.2 | 0.7 | 1.2 | Polychaeta | 0.2 | 0.2 | 99.4 | 0.4 | 2.1 |
| Bivalvia | 0.3 | 0.4 | 98.8 | 0.1 | 0.7 | Polychaeta | 0.2 | 0.2 | 99.5 | 0.0 | 2.1 | Bivalvia | 0.2 | 0.2 | 99.6 | 0.7 | 0.5 |
| Cladocera | 0.3 | 0.4 | 99.2 | 0.1 | 0.1 | Halacaroidea | 0.1 | 0.2 | 99.6 | 0.8 | 0.6 | Amphipoda | 0.1 | 0.1 | 99.8 | 0.0 | 1.2 |
| Pycnogonida | 0.2 | 0.2 | 99.4 | 0.0 | 0.3 | Bivalvia | 0.1 | 0.2 | 99.8 | 0.1 | 0.5 | Insecta | 0.1 | 0.1 | 99.9 | 0.0 | 0.4 |
| Oligochaeta | 0.2 | 0,.2 | 99.6 | 0.8 | 0.3 | Insecta | 0.1 | 0.1 | 99.9 | 0.0 | 0.4 | Pycnogonida | 0.0 | 0.1 | 99.9 | 0.3 | 0.0 |
| Kinorhyncha | 0.1 | 0.2 | 99.8 | 0.1 | 0.0 | nectochaetes | 0.0 | 0.0 | 99.9 | 0.0 | 0.3 | nectochaetes | 0.0 | 0.0 | 100.0 | 0.0 | 0.3 |
| Isopoda | 0.1 | 0.1 | 99.9 | 0.2 | 0.0 | Isopoda | 0.0 | 0.0 | 99.9 | 0.2 | 0.0 | Cladocera | 0.0 | 0.0 | 100.0 | 0.1 | 0.0 |
| Polychaeta | 0.1 | 0.1 | 100.0 | 0.0 | 0.4 | Cladocera | 0.0 | 0.0 | 100.0 | 0.1 | 0.0 | Nemertina | 0.0 | 0.0 | 100.0 | 0.0 | 0.1 |
| Nemertina | 0.0 | 0.0 | 100.0 | 0.0 | 0.0 | Kinorhyncha | 0.0 | 0.0 | 100.0 | 0.1 | 0.0 | Tanaidacea | 0.0 | 0.0 | 100.0 | 0.0 | 0.1 |
| Insecta | 0.0 | 0.0 | 100.0 | 0.0 | 0.0 | Nemertina | 0.0 | 0.0 | 100.0 | 0.0 | 0.1 | Kinorhyncha | 0.0 | 0.0 | 100.0 | 0.0 | 0.0 |
| nectochaetes | 0.0 | 0.0 | 100.0 | 0.0 | 0.0 | Pycnogonida | 0.0 | 0.0 | 100.0 | 0.0 | 0.0 | Isopoda | 0.0 | 0.0 | 100.0 | 0.0 | 0.0 |
Table 4.
Results of two-way PERMANOVA testing differences among sites for the total meiofaunal abundance, number of taxa, diversity, evenness, and Ne/Co ratio (Euclidean distance based). Significant p values are marked, * for p < 0.05; ** for p < 0.01.
Table 4.
Results of two-way PERMANOVA testing differences among sites for the total meiofaunal abundance, number of taxa, diversity, evenness, and Ne/Co ratio (Euclidean distance based). Significant p values are marked, * for p < 0.05; ** for p < 0.01.
| Variable | Factors | Total Sum of Squares | df | Mean Square | Pseudo-F | p | Pairwise |
|---|---|---|---|---|---|---|---|
| Total abundance | Sites | 0.54 | 2 | 0.27 | 10.74 | 0.0003 ** | C1 vs. C3 p = 0.01 *; C2 vs. C3 p < 0.01 ** |
| Periods | 0.01 | 1 | 0.01 | 0.46 | 0.57 | ||
| Sites × Periods | 0.11 | 2 | 0.06 | 2.2 | 0.11 | ||
| Residual | 0.75 | 30 | 0.03 | ||||
| Total | 1.41 | 35 | |||||
| number of taxa | Sites | 0.49 | 2 | 0.24 | 6.15 | 0.0023 ** | C2 vs. C3 p < 0.05 * |
| Periods | 0.03 | 1 | 0.03 | 0.76 | 0.42 | ||
| Sites × Periods | 0.02 | 2 | 0.01 | 0.21 | 0.76 | ||
| Residual | 1.19 | 30 | 0.04 | ||||
| Total | 1.73 | 35 | |||||
| Shannon-diversity | Sites | 0.08 | 2 | 0.04 | 0.64 | 0.8 | - |
| Periods | 0.07 | 1 | 0.07 | 1.11 | 0.31 | ||
| Sites × Periods | 0.16 | 2 | 0.08 | 1.29 | 0.22 | ||
| Residual | 1.9 | 30 | 0.06 | ||||
| Total | 2.22 | 35 | |||||
| Pielou-evenness | Sites | 0.31 | 2 | 0.16 | 2.52 | 0.0166 * | C1 vs. C3 p < 0.05 * |
| Periods | 0.17 | 1 | 0.17 | 2.71 | 0.0356 * | ||
| Sites × Periods | 0.19 | 2 | 0.1 | 1.56 | 0.13 | ||
| Residual | 1.86 | 30 | 0.06 | ||||
| Total | 2.53 | 35 | |||||
| Ne/Co ratio | Sites | 1.16 | 2 | 0.58 | 3 | 0.02 * | C1 vs. C3 p < 0.01 ** |
| Periods | 0.24 | 1 | 0.24 | 1.25 | 0.28 | ||
| Sites × Periods | 0.71 | 2 | 0.36 | 1.83 | 0.13 | ||
| Residual | 5.81 | 30 | 0.19 | ||||
| Total | 7.92 | 35 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Semprucci, F.; Gravina, M.F.; Magni, P. Meiofaunal Dynamics and Heterogeneity along Salinity and Trophic Gradients in a Mediterranean Transitional System. Water 2019, 11, 1488. https://doi.org/10.3390/w11071488
AMA Style
Semprucci F, Gravina MF, Magni P. Meiofaunal Dynamics and Heterogeneity along Salinity and Trophic Gradients in a Mediterranean Transitional System. Water. 2019; 11(7):1488. https://doi.org/10.3390/w11071488
Chicago/Turabian StyleSemprucci, Federica, Maria Flavia Gravina, and Paolo Magni. 2019. "Meiofaunal Dynamics and Heterogeneity along Salinity and Trophic Gradients in a Mediterranean Transitional System" Water 11, no. 7: 1488. https://doi.org/10.3390/w11071488
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.