Flow Cytometric Analysis of Freshwater Cyanobacteria: A Case Study


Department of Biological Sciences, Seton Hall University, NJ 07079, USA
*
Author to whom correspondence should be addressed.
Water 2019, 11(7), 1422; https://doi.org/10.3390/w11071422
Submission received: 1 June 2019
/
Revised: 3 July 2019
/
Accepted: 9 July 2019
/
Published: 11 July 2019
(This article belongs to the Section Water Quality and Contamination)
Abstract
:Eutrophication is a process that occurs due to the excessive accumulation of nutrients, primarily nitrogen and phosphorus, from natural and anthropogenic sources. This phenomenon causes cyanobacterial overgrowth, which over time leads to cyanobacterial harmful algal blooms (CHABs) that affect public drinking water sources and water sites with recreational usage. The rapid detection of bloom-forming cyanobacteria in freshwater bodies is critical in order to implement prevention strategies. Cyanobacteria contain phycobiliproteins such as phycoerythrin and allophycocyanin as part of the phycobilisome that allows autofluorescence. In this study, samples from 36 freshwater bodies in 14 New Jersey counties were collected and analyzed using flow cytometry with forward-scatter phycoerythrin and allophycocyanin parameters. Pure cultures of Synechococcus sp. IU 625, Cylindrospermum spp. and Microcystis aeruginosa were used as references. The results revealed that 17 out of the 36 analyzed sites contained all three references and related species. Seven sites showed Microcystis and Cylindrospermum-like species, while four sites indicated Microcystis and Cylindrospermum-like species. Six water bodies showed Cylindrospermum-like species, and two sites showed Microcystis-like species. Polymerase chain reaction (PCR)-based assays further confirmed the flow cytometric results. The findings from this study suggest that flow cytometry could potentially serve as a rapid method for freshwater cyanobacteria detection and screening.
1. Introduction
Cyanobacteria, also known as blue green algae, are some of the earliest dated oxygen-releasing prokaryotes, originating about 3.5 billion years ago [1]. Their size ranges from 0.5 to 50+ µm, and they may be composed of different cellular arrangements, such as unicellular Synechococcus spp., filamentous Cylindrospermum spp., and colonial Microcystis spp. Cyanobacteria can be found in marine and freshwater environments, including lakes, ponds, rivers, brackish water and water reservoirs. Moreover, these bacteria can be terrestrial, colonize in volcano ashes, and even colonize in the human gut. Cyanobacteria are able to survive and outcompete other biological species in eutrophied water bodies and, consequently, can over-grow nutrients such as nitrogen and phosphorus, leading to the formation of algal blooms. Climate change as well as human disruptions such as agriculture, industrial and recreational activities, and sewage effluent all lead to eutrophication [2]. The bloom masses may cover the water surface, lowering the dissolved oxygen and causing hypoxia in the water. Although not all cyanobacteria are toxic, many possess the ability to release toxins and produce secondary metabolites known as cyanotoxins, including hepatotoxins, neurotoxins and dermatoxins, which are toxic to other cells, organisms, fish, animals and humans. Cyanobacterial harmful algal blooms (CHABs) are considered a significant concern to the ecosystem. In addition, their toxins can be very dangerous to the public, who utilize the water for drinking and recreational purposes. Cyanobacterial toxicity can vary from one bloom to another, where specific species can secrete a certain toxin or even multiple species can release different cyanotoxins [3,4]. Intracellular toxins for most cyanobacteria are not secreted unless an environmental stress factor induces cell lysis [4,5].
Approximately 80% of human exposure to cyanotoxins is from the ingestion of contaminated water [6,7]. The United States Environmental Protection Agency (USEPA) lists several cyanotoxins on the candidate list of contaminates that primarily affect public drinking water, including microcystin, cylindrospermopsin and the anatoxin-a group. Microcystins, primarily found in fresh and brackish water, causes toxicity to the liver by triggering liver inflammation and/or liver hemorrhage due to cytoskeletal damage induced by inhibition of serine/threonine-specific protein phosphatases [8,9]. Anatoxin-a involves the binding of its toxin to a neuronal pre-synaptic acetylcholine receptor, where nicotine receptors are mimicked, thus, causing neurological effects such as paralysis and even death from respiratory arrest [10,11,12,13,14,15]. Other cyanobacterial neurotoxins include saxitoxin (also known as paralytic shellfish toxin) and non-proteinous amino acid ß-methylamino-L-alanine (BMAA) [16,17,18,19]. Cylindrospermopsin (CYN), a cytotoxin with five variants or analogues found further below the water surface, can have its toxin released extracellularly in up to 90% of the total CYN content in the water column [4,13,20,21,22,23,24,25]. The toxic mechanism of cylindrospermopsin involves protein synthesis inhibition, which leads to kidney and liver failure [9]. It can also induce oxidative stress and can cause cell-cycle arrest, apoptosis or necrosis in various cell lines [26]. The World Health Organization (WHO) has established a recommended limit for microcystin-LR of 1.0 µg/L in drinking water. Most countries follow this WHO guideline, but some have minor variations that can range from 1.0 to 1.5 µg/L [27]. Children are at greater risk due to their body size to weight ratio. USEPA has provided separate guideline limits for children younger than six years old of 0.3 and 0.7 µg/L for microcystin and cylindrospermopsin, respectively [12,28].
Current treatments for algal bloom include the use of algaecides and chlorine, but they have been shown to cause cell lysis and, subsequently, the secretion of toxins [29,30]. Furthermore, while microfiltration and ultrafiltration have been shown to be effective for the prevention of intracellular toxins with up to 98% removal of Microcystis aeruginosa, they are ineffective for the removal of extracellular toxins [13,31]. Recently, researchers have looked at using cyanophages, host-specific bacteriophages that have the ability to infect species-specific cyanobacteria, as a possible biological prevention approach for cyanobacterial blooms. Further knowledge of cyanophages can aid in the reduction of bloom-forming cyanobacteria and increase water quality and the health of the ecosystem [32].
Finally, there is an immediate need for rapid detection of harmful algal blooms worldwide in order to avoid and or halt further issues and reduce the need for chemical treatments that ultimately cause cell lysis and toxin secretion in water bodies. Currently, implemented detection methods contain some limitations. For instance, phenotypic identification through microscopy can be time-consuming, especially with high volumes of samples for monitoring purposes. Molecular identifications such as polymerase chain reaction (PCR) are more accurate in terms of specificity and sensitivity; however, they are not cost effective and cannot detect single cells [33]. Moreover, biochemical and immunoassays can be utilized, but require a field expert, expensive equipment, and may result in false positives [34]. Flow cytometry analysis may be an effective tool for analyzing routine environmental water samples for the prevention of algal blooms [31]. Cyanobacteria contain phycobillisomes, which are composed of autofluorescent pigments known as phycoerythrin (PE), phycocyanin (PC), and allophycocyanin (APC) [35,36]. Flow cytometry detects these natural pigments in photosynthetic microorganisms, including PE and APC [31,36]. Previous reports have discussed the utilization of flow cytometry to study cyanobacteria and monitor microbes in drinking water sources [37,38]. This study aims to determine the cyanobacterial profiles of freshwater bodies by comparing water samples to the reference cyanobacteria with flow cytometry. Synechococcus sp. IU 625, a unicellular cyanobacterium, Microcystis aeruginosa, and Cylindrospermum spp. have been commonly used as cyanobacterial algal bloom indicators [4,11] and are included in this study as reference cyanobacteria.
2. Materials and Methods
2.1. Water Sample Collection and Processing
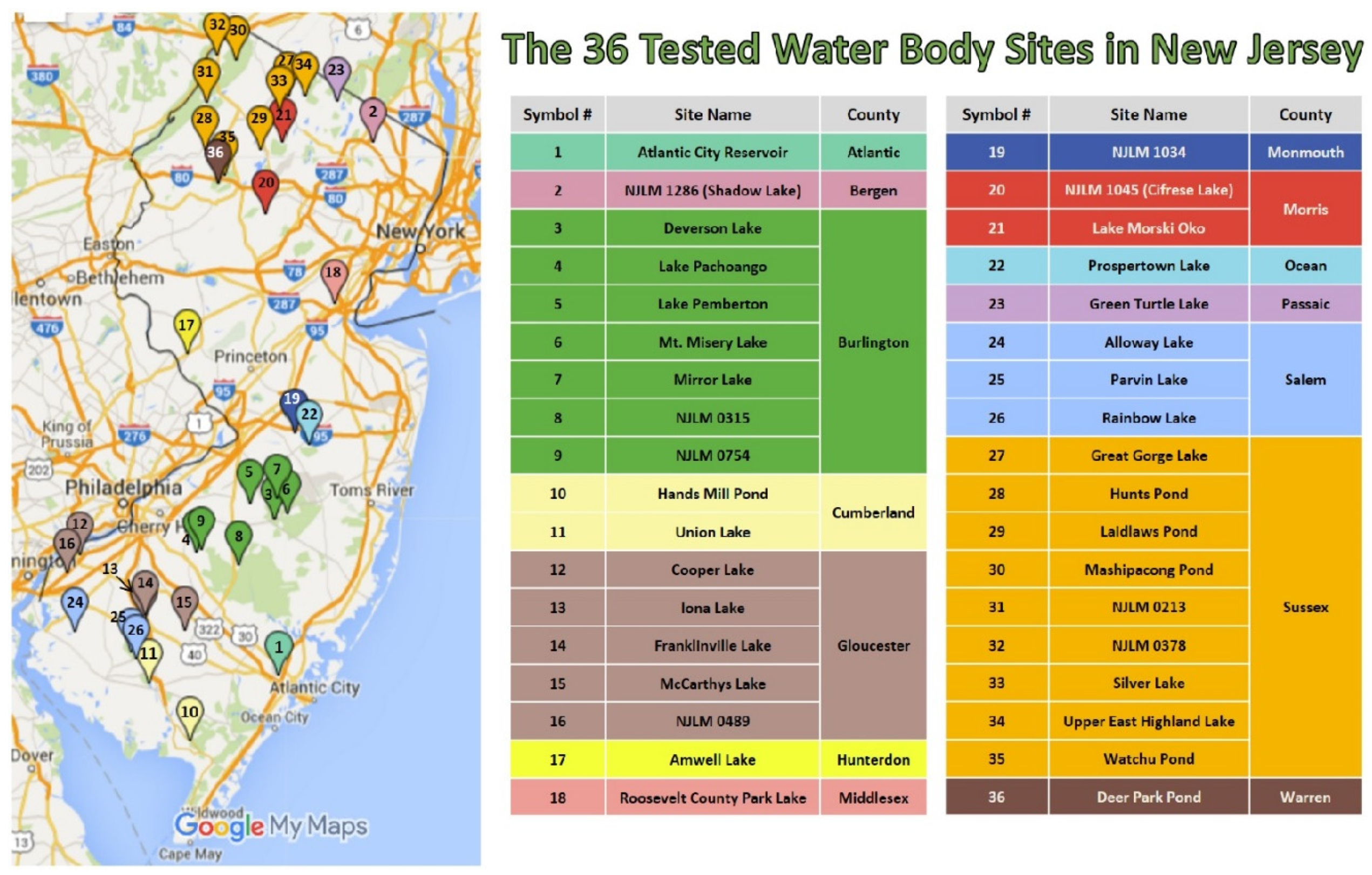
Water samples from 36 water bodies (250 mL per site in Nalgene bottles) in New Jersey (Figure 1) were obtained by the New Jersey Department of Environmental Protection (NJDEP) in summer (May to August), 2015. These water bodies are utilized for recreational purposes, drinking water, or water supply reservoirs. The Global Positioning System (GPS) coordinates as well as water temperature (°C), pH and dissolved oxygen (mg/L) were measured and recorded on site. Water samples were collected one meter below the surface.
2.2. Filtration
A Thermo Scientific™ Nalgene™ (Thermo Scientific, Rochester, NY, USA) vacuum filtration unit with polycarbonate membrane filters (Sterlitech Corporation, Kent, WA, USA) was used for filtration. The water samples were thoroughly mixed and serial filtered through 30 and 0.4 μm membrane filters, sequentially.
2.3. Flow Cytometry
Flow cytometric assays were conducted using a MACSQuant® Analyzer, calibrated with MacsQuant calibration beads (Miltenyi Biotec, Inc., SanDiego, CA, USA). The three standard reference cultures Microcystis, S. IU 625 and Cylindrospermum spp. with Optical Density (OD750nm) of 0.591, 0.521 and 0.640, respectively, were run through the flow cytometer at fixed voltage settings with various dilutions. The water samples for each site and the <30 µm filtrates were run through the flow cytometer, with the same voltage settings as the standards. The low flow rate and mix sample options were selected for this analysis. The voltage channels for forward scatter (FSC), side scatter (SSC), B2 phycoerythrin (PE), R1 allophycocyanin (APC) were 250 V, 200 V, 800 V, and 275 V, respectively. Each uptake contained 100 μL of the sample. The analysis was carried out using FSC and PE gating strategy with the software FlowJo vX.0.7 (Tree Star, Inc., Ashland, OR, USA).
2.4. Molecular Assays
Chelex DNA extraction was conducted for all collected water samples. Polymerase chain reaction (PCR) was carried out with CYA359F/CYA781R, the primer set that amplifies cyanobacterial 16S rRNA genes [39], to confirm the flow cytometry results. MSF/MSR, a primer set to detect the presence of Microcystin Synthetase A Gene (mcyA) in the Cyanobacterial Genus Microcystis was also included [40]. A 1% agarose gel electrophoresis was used to visualize the PCR products. PCR products were subsequently sequenced with an ABI PRISM® 310 Genetic Analyzer (Applied Biosystems, Foster City, CA, USA). Multiple Sequence Comparison by Log-Expectation (MUSCLE) 3.8 was utilized to analyze the 16S rRNA genes of Cylindrospermum spp., Microcystis spp., and Synechococcus spp. [41].
3. Results
3.1. Water Collection Sites
Water properties were recorded for all 36 sites (Table 1). Among all the water samples collected and tested, the water temperature ranged from 16.97 (NJLM 0213) to 30.9 °C (Alloway Lake). The pH ranged from 4.09 (Mt. Misery Lake) to 9.75 (Cooper Lake), with the majority of sites between 6 and 9. The dissolved oxygen ranged from 1.66 (NJLM 0213) to 19.1 mg/L (Cooper Lake).
3.2. Filtration Cytometric Analysis of Three Reference Cyanobacteria
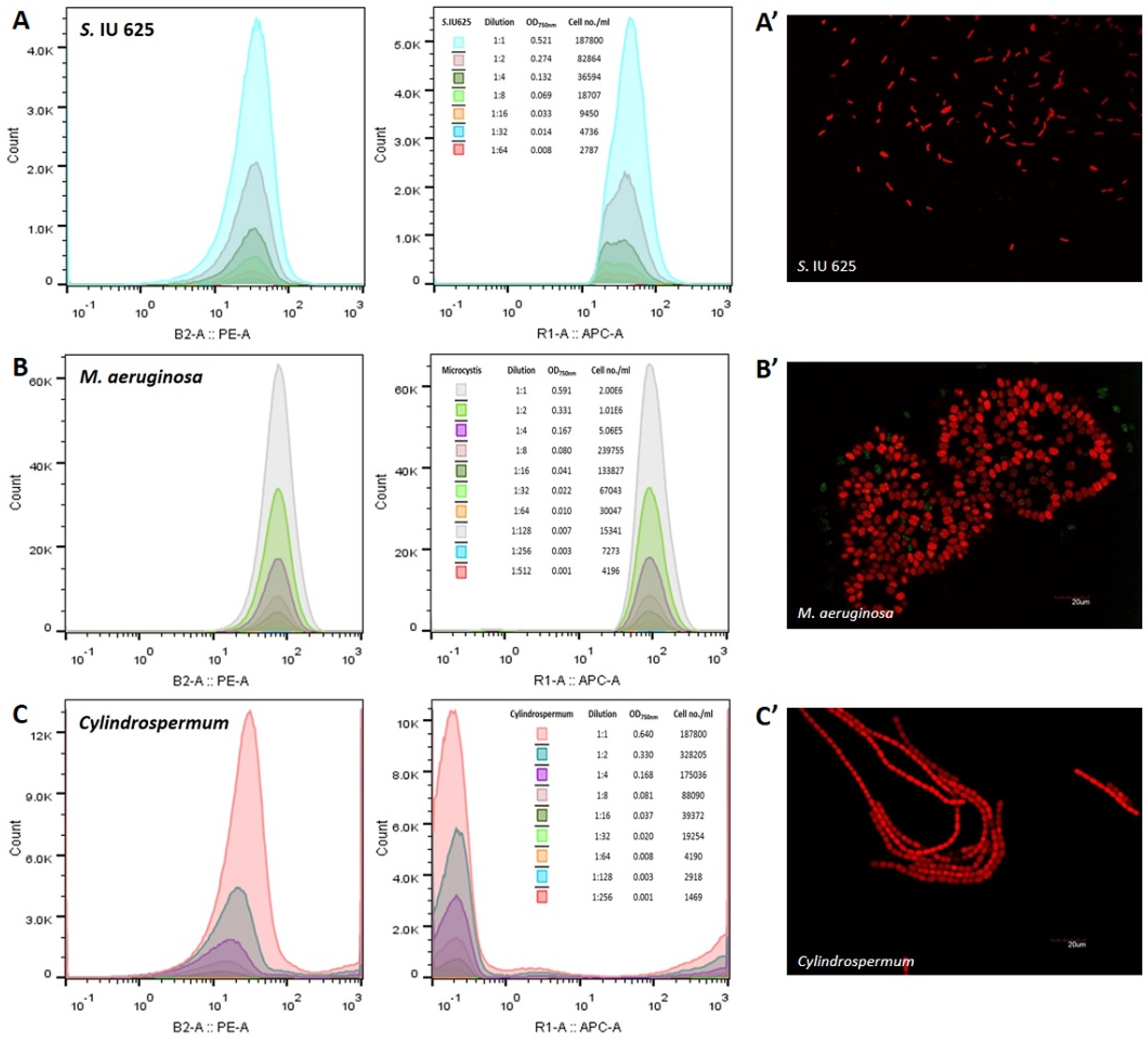
Three reference species, namely, Synechococcus sp. IU 625, Microcystis aeruginosa, and Cylindrospermum spp., were measured and analyzed through flow cytometry. The results revealed that M. aeruginosa is detectable at an accurate range from an OD750nm of 0.001 and 4.196 × 103 cells/mL up to an OD750nm of 0.591 and 2.0 × 106 cells/mL, based on the phycoerythrin (PE) and allophycocyanin (APC) parameters (Figure 2). Synechococcus sp. IU 625 can be accurately detected at an OD750nm range of 0.008 to 0.521 and cell count of 2.787 × 103 to 1.878 × 105 cells/mL (Figure 2). The detectable range of Cylindrospermum spp. spans from an OD750nm of 0.001 to 0.640 and cell count of 1.479 × 103 to 7.21 × 105 cells/mL (Figure 2). M. aeruginosa contains higher PE and APC intensities when compared with S. IU 625 and Cylindrospermum spp.
3.3. Flow Cytometric Analysis of 36 Water Bodies in New Jersey
Samples from a total of 36 water bodies across NJ were collected and analyzed using flow cytometric assays. Three pure cyanobacterial strains, Cylindrospermum spp., Microcystis aeruginosa and Synechococcus sp. IU 625, were included in this analysis. Using a color-gating algorithm with the forward-scatter (FSC) versus phycoerythrin (PE) parameters, the reference cyanobacteria were shown in three populations: Cylindrospermum spp. in red, Microcystis aeruginosa in orange, and Synechococcus sp. IU 625 in blue color (Figure 3).
3.3.1. Microcystis spp., Synechococcus sp. IU 625 and Cylindrospermum spp. Related Species
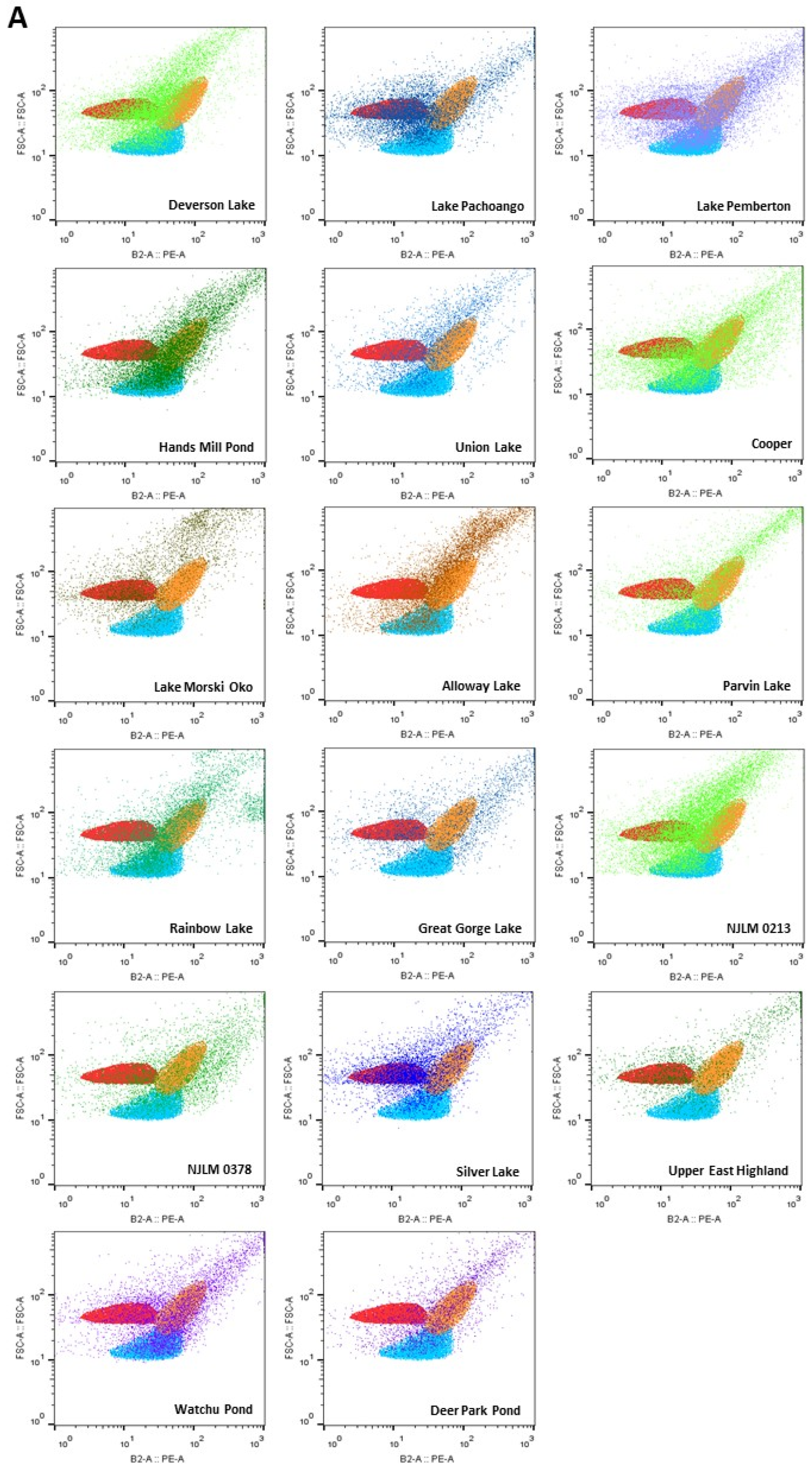
Flow cytometric analyses showed that 17 water bodies, including Deverson Lake, Lake Pachoango, Lake Pemberton, Cooper Lake, Hands Mill, Lake Morski Oko, Alloway Lake, Parvin Lake, Rainbow Lake, Union Lake, Great Gorge Lake, NJLM 0213, NJLM 0378, Silver Lake, Upper East Highland, Watchu Pond, and Deer Park Pond, contained cyanobacteria including Microcystis spp. (orange), Synechococcus spp. IU 625 (blue), Cylindrospermum spp. (red), and their related species based on the FSC vs. PE profiles (Figure 3A).
3.3.2. Microcystis spp. and Synechococcus sp. IU 625 Related Species
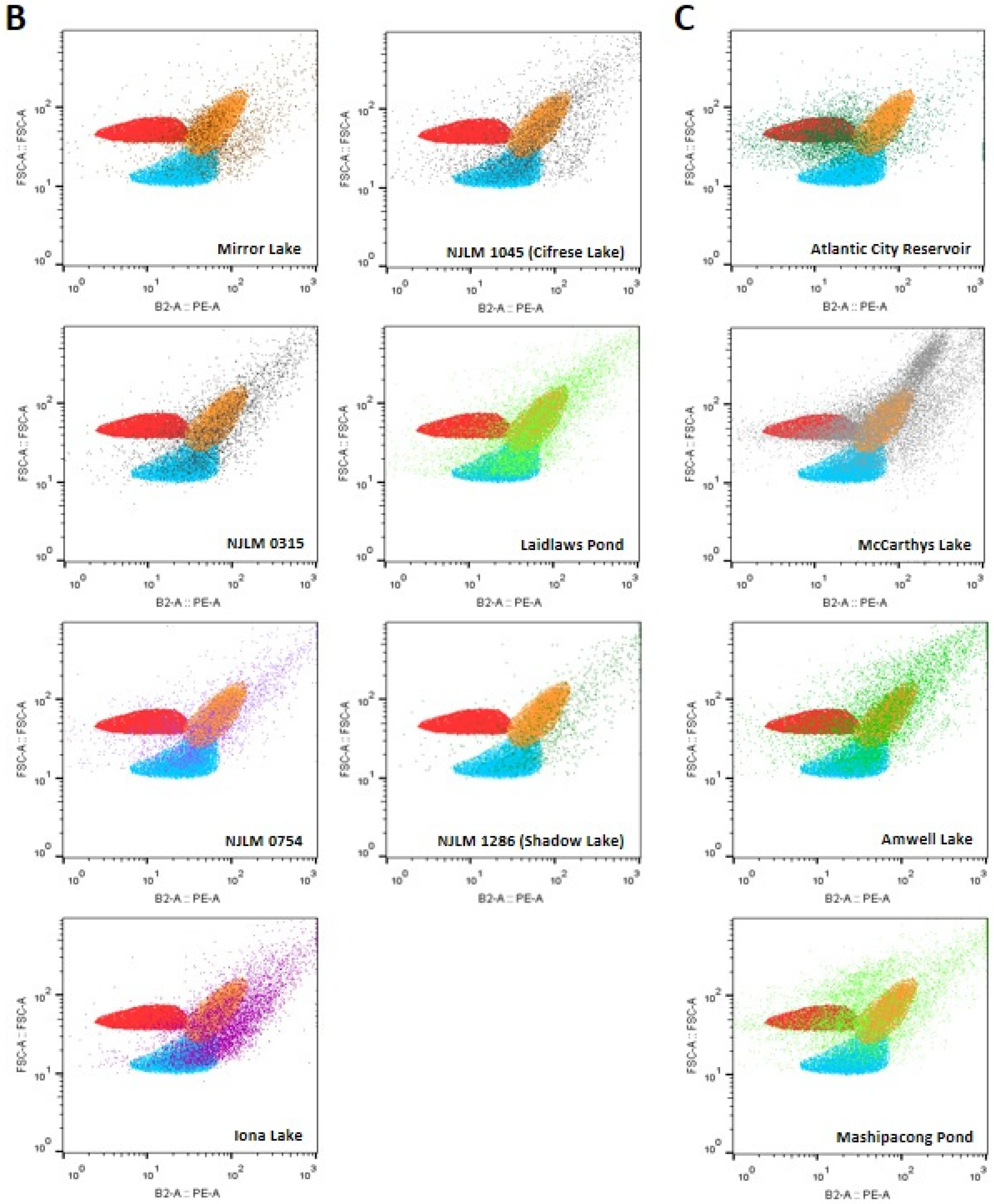
For seven out of the 36 water bodies, including NJLM 1286 (Shadow Lake), Mirror Lake, NJLM 0315, Iona Lake, NJLM 1045 (Cifrese Lake), Laidlaw Pond, and NJLM 0754, it was revealed that the phytoplanktons in these sites contained cyanobacteria of mostly Microcystis spp. (orange) and Synechococcus sp. IU 625 (blue) related species, based on the FSC vs. PE profiles (Figure 3B).
3.3.3. Microcystis spp., and Cylindrospermum spp. Related Species
For four out of the 36 water bodies, including Atlantic City Reservoir, McCarthys Lake, Amwell Lake, and Mashipacong Pond, it was shown that the phytoplankton in these sites contained mostly Microcystis spp. (orange) and Cylindrospermum spp. (red) related species, based on the FSC vs. PE profiles (Figure 3C).
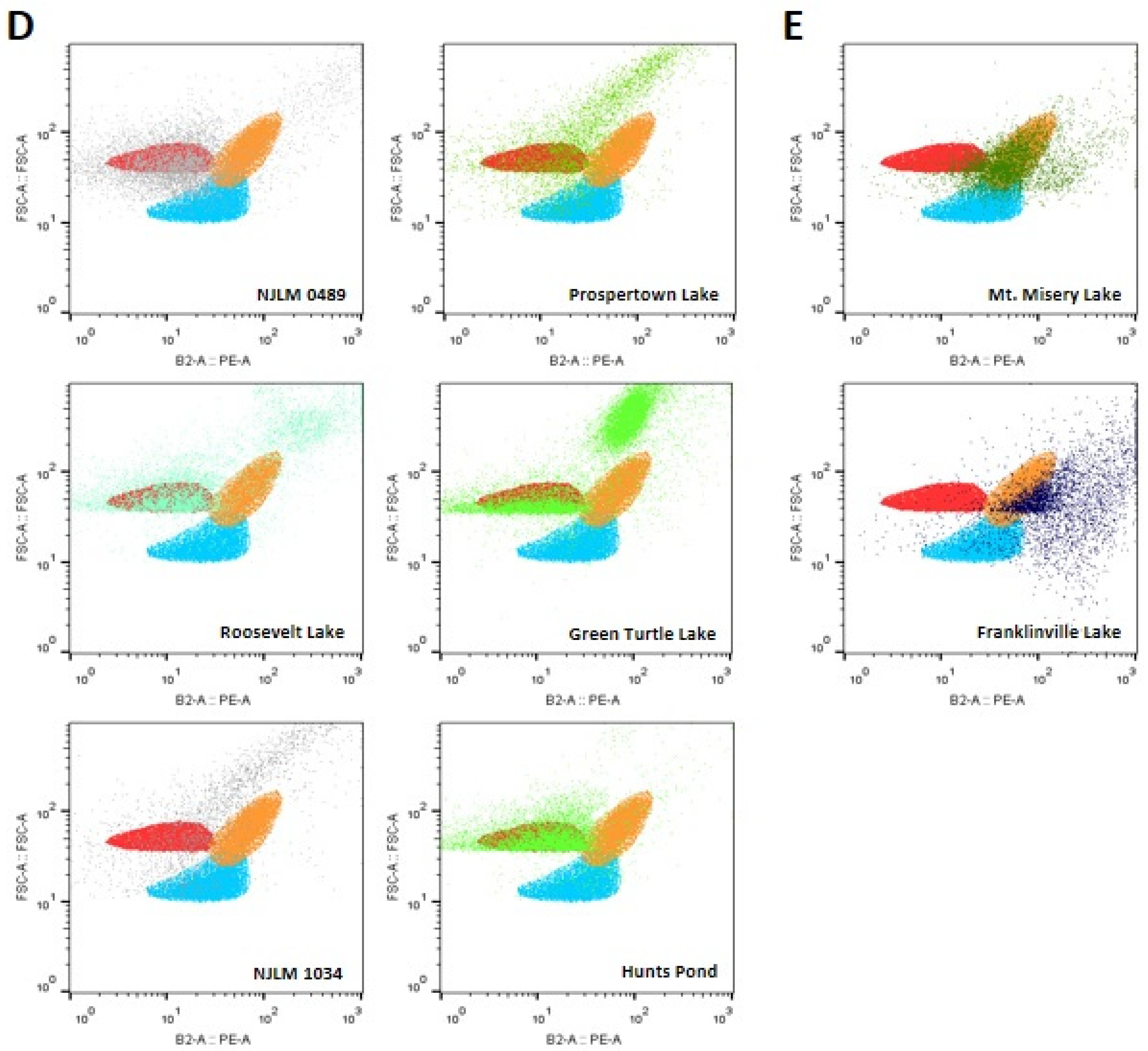
3.3.4. Cylindrospermum spp. Related Species
Six out of the 36 water bodies, including NJLM 0489, Roosevelt Lake, and Hunts Pond, revealed primarily Cylindrospermum spp. (red) related species (Figure 3D). The results also showed that the cyanobacteria in NJLM 1034, Prospertown Lake, and Green Turtle Lake water bodies contained mainly Cylindrospermum spp. (red) related species and an undetermined population, based on the FSC vs. PE profiles (Figure 3D).
3.3.5. Microcystis spp. and Related Species
The results showed that Mt. Misery and Franklinville Lake contained cyanobacteria of mostly Microcystis spp. (orange) related species, based on the FSC vs. PE profiles (Figure 3E). Franklinville Lake water samples showed another phycoerythrin-rich population.
3.4. Molecular Analysis of Cyanobacteria
4. Discussion
In this study, water samples from 36 water bodies throughout 14 New Jersey counties were analyzed via flow cytometry. The water chemistry, including water temperature (°C), pH, and dissolved oxygen, was recorded by NJDEP, as seen in Table 1. Among all sites, NJLM 0213 contained the lowest water temperature and dissolved oxygen, with 16.97 °C and 1.66 mg/L, respectively. The highest water temperature was 30.9 °C from Alloway Lake. The highest pH and dissolved oxygen was found in Cooper Lake, with 9.75 and 19.1 mg/L, respectively. The lowest pH was 4.09 from Mt. Misery Lake. The optimal pH range for cyanobacteria is between 6 and 9 [4]. Most of the sites analyzed fall within that range, allowing cyanobacteria to grow and possibly accumulate (Table 1). Dissolved oxygen is a water property that is crucial where there are concentrations of less than 4 mg/L, which can be considered hypoxic to the water ecosystem [43,44]. Four sites from Sussex County, NJLM 0213, NJLM 0378, Silver Lake and Watchu Pond, contained very little dissolved oxygen, with 1.66, 2.65, 3.32 and 3.69 mg/L, respectively (Table 1), and showed the presence of all three reference Microcystis spp., Synechococcus sp. IU 625, Cylindrospermum spp. and related species as expected (Figure 3A). All three cyanobacterial reference species were found in wide ranges of temperature (16.97 °C to 30.9 °C), pH (4.44 to 9.75) and dissolved oxygen (1.66 to 19.17 mg/L). Microcystis spp. was detected in the environment with pH as low as 4.09.
The water profile of each of the 36 New Jersey water sites was obtained via flow cytometric analysis in about 4 to 5 min, per sample. Thus, this study further confirmed that flow cytometry can be used to rapidly process, screen and accurately detect a large volume of environmental water samples. The profile of different cyanobacteria species can be directly detected in real time without staining or lysis, due to cyanobacterial autofluorescence. Thus, flow cytometry allows early preventive measurements to be implemented and allows for a more cost- and time-efficient routine sampling method. As seen in Figure 3, cyanobacteria were successfully detected in all 36 collected water sites when analyzed by flow cytometry. Most of the cyanobacteria or phytoplankton species were mainly phycoerythrin-rich. It is important to note that reference cyanobacterial cultures were maintained in optimal conditions, which may cause them to exhibit different pigment levels when compared with the same species in lakes. Furthermore, PCR-based assays were conducted as a complementary method to confirm the flow cytometric analysis results [42,45]. The primers utilized were to amplify 16S rRNA genes for each reference cyanobacterium [39]. A microcystin synthetase primer was also included for cyanotoxin detection. The results from PCR-based assays indicated the presence of three reference cyanobacteria in all sites, but no microcystin was detected in any of the testing water samples.
Finally, this study highlighted the use of flow cytometry for the rapid detection of cyanobacteria species in 36 New Jersey water bodies. Flow cytometry is an efficient, rapid routine water sample testing method [46], so a larger volume sampling of freshwater bodies can be achieved. A flow cytometer with sorting capability could further isolate various phytoplankton taxa from mixed environmental samples in water bodies [47]. In addition, portable flow cytometers are available for the samples to be processed on site for more accurate analyses. In summary, flow cytometry could be used as an accompanying method for the rapid detection of cyanobacteria.
Author Contributions
Conceptualization, T.C.; methodology, T.C.; investigation, R.P.; microscopy, R.N.J.; formal analysis, R.P., and T.C.; resources, T.C.; data curation, T.C.; writing—original draft preparation, R.P., A.d.O. and T.C.; writing—review and editing, R.P., A.d.O. and T.C.; visualization, R.P., R.N.J., and T.C.; supervision, T.C.; project administration, T.C.; funding acquisition, T.C.
Funding
This research was funded by Seton Hall University (SHU) Biological Sciences Department Annual Research Fund and William and Doreen Wong Foundation to T.C., and SHU Graduate Teaching Assistantship in the Department of Biological Sciences to A.O. and R.N.
Acknowledgments
The authors would like to thank Johannus Franken, Brian Taylor, Yun-Shen Lee and the rest of NJDEP for taking the time to collect and transport the samples from the NJ water bodies.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Schopf, J.W. The Fossil Record: Tracing the Roots of the Cyanobacterial Lineage. In The Ecology of Cyanobacteria; Whitton, B.A., Potts, M., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 13–35. [Google Scholar]
- Paerl, H.W. Mitigating harmful cyanobacterial blooms in a human- and climatically-impacted world. Life 2014, 4, 988–1012. [Google Scholar] [CrossRef] [PubMed]
- Funari, E.; Testai, E. Human health risk assessment related to cyanotoxins exposure. Crit. Rev. Toxicol. 2008, 38, 97–125. [Google Scholar] [CrossRef] [PubMed]
- D’Anglada, L.V.; Donohue, J.M.; Strong, J. Health Effects Support Document for the Cyanobacterial Toxin Microcystins; USEPA: Washington, DC, USA, 2015.
- Masten, S.C.B. Cylindrospermopsin—Review of Toxicological Literature; Toxicological Summary for Cylindrospermopsin; Final Report 2000; National Institute of Environmental Health Sciences Research: Triangle Park, NC, USA, 2001.
- World Health Organization. Guidelines for drinking-water quality. In Health Criteria and Other Supporting Information: Addendum; World Health Organization: Geneva, Switzerland, 1998; Volume 2. [Google Scholar]
- Merel, S.; Walker, D.; Chicana, R.; Snyder, S.; Baures, E.; Thomas, O. State of knowledge and concerns on cyanobacterial blooms and cyanotoxins. Environ. Int. 2013, 59, 303–327. [Google Scholar] [CrossRef] [PubMed]
- Jungblut, A.D.; Neilan, B.A. Molecular identification and evolution of the cyclic peptide hepatotoxins, microcystin and nodularin, synthetase genes in three orders of cyanobacteria. Arch. Microbiol. 2006, 185, 107–114. [Google Scholar] [CrossRef] [PubMed]
- van Apeldoorn, M.E.; van Egmond, H.P.; Speijers, G.J.; Bakker, G.J. Toxins of cyanobacteria. Mol. Nutr. Food Res. 2007, 51, 7–60. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, W.W. Cyanobacteria secondary metabolites—The cyanotoxins. J. Appl. Bacteriol. 1992, 72, 445–459. [Google Scholar] [CrossRef] [PubMed]
- Chorus, I.; Bartram, J. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; E & FN Spon on behalf of the World Health Organization: London, UK, 1999; 416p. [Google Scholar]
- Farrer, D.; Counter, M.; Hillwig, R.; Cude, C. Health-based cyanotoxin guideline values allow for cyanotoxin-based monitoring and efficient public health response to cyanobacterial blooms. Toxins 2015, 7, 457–477. [Google Scholar] [CrossRef]
- USEPA. Cyanobacteria and Cyanotoxins: Information for Drinking Water Systems; USEPA: Washington, DC, USA, 2014.
- USEPA. Health Effects Support Document for the Cyanobacterial Toxin Anatoxin-a; USEPA: Washington, DC, USA, 2015.
- Wonnacott, S.; Gallagher, T. The Chemistry and Pharmacology of Anatoxin-a and Related Homotropanes with respect to Nicotinic Acetylcholine Receptors. Mar. Drugs 2006, 4, 228–254. [Google Scholar] [CrossRef] [Green Version]
- Jakubowska, N.; Szelag-Wasielewska, E. Toxic picoplanktonic cyanobacteria—Review. Mar. Drugs 2015, 13, 1497–1518. [Google Scholar] [CrossRef]
- Wiese, M.; D’Agostino, P.M.; Mihali, T.K.; Moffitt, M.C.; Neilan, B.A. Neurotoxic alkaloids: Saxitoxin and its analogs. Mar. Drugs 2010, 8, 2185–2211. [Google Scholar] [CrossRef]
- Chiu, A.S.; Gehringer, M.M.; Welch, J.H.; Neilan, B.A. Does alpha-amino-beta-methylaminopropionic acid (BMAA) play a role in neurodegeneration? Int. J. Environ. Res. Public Health 2011, 8, 3728–3746. [Google Scholar] [CrossRef] [PubMed]
- Chiu, A.S.; Gehringer, M.M.; Braidy, N.; Guillemin, G.J.; Welch, J.H.; Neilan, B.A. Excitotoxic potential of the cyanotoxin beta-methyl-amino-L-alanine (BMAA) in primary human neurons. Toxicon 2012, 60, 1159–1165. [Google Scholar] [CrossRef] [PubMed]
- Chiswell, R.K.; Shaw, G.R.; Eaglesham, G.; Smith, M.J.; Norris, R.L.; Seawright, A.A.; Moore, M.R. Stability of cylindrospermopsin, the toxin from cyanobacterium, Cylindropsermopsis raciborskii: Effect of pH, temperature, and sunlight on decomposition. Environ. Toxicol. 1999, 14, 155–161. [Google Scholar] [CrossRef]
- USEPA. Health Effects Support Document for the Cyanobacterial Toxin Cylindrospermopsin; USEPA: Washington, DC, USA, 2015.
- Cartmell, C.; Evans, D.M.; Elwood, J.M.L.; Fituri, H.S.; Murphy, P.J.; Caspari, T.; Poniedzialek, B.; Rzymski, P. Synthetic analogues of cyanobacterial alkaloid cylindrospermopsin and their toxicological activity. Toxicol. In Vitro 2017, 44, 172–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poniedzialek, B.; Rzymski, P.; Kokocinski, M. Cylindrospermopsin: Water-linked potential threat to human health in Europe. Environ. Toxicol. Pharmacol. 2012, 34, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Bormans, M.; Lengronne, M.; Brient, L.; Duval, C. Cylindrospermopsin accumulation and release by the benthic cyanobacterium Oscillatoria sp. PCC 6506 under different light conditions and growth phases. Bull. Environ. Contam. Toxicol. 2014, 92, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Rucker, J.; Stuken, A.; Nixdorf, B.; Fastner, J.; Chorus, I.; Wiedner, C. Concentrations of particulate and dissolved cylindrospermopsin in 21 Aphanizomenon-dominated temperate lakes. Toxicon 2007, 50, 800–809. [Google Scholar] [CrossRef] [PubMed]
- Poniedzialek, B.; Rzymski, P.; Karczewski, J. The role of the enzymatic antioxidant system in cylindrospermopsin-induced toxicity in human lymphocytes. Toxicol. In Vitro 2015, 29, 926–932. [Google Scholar] [CrossRef] [PubMed]
- Ibelings, B.W.; Backer, L.C.; Kardinaal, W.E.; Chorus, I. Current approaches to cyanotoxin risk assessment and risk management around the globe. Harmful Algae 2015, 49, 63–74. [Google Scholar] [CrossRef] [PubMed]
- USEPA. Drinking Water Health Advisories for Two Cyanobacterial Toxins; USEPA: Washington, DC, USA, 2015.
- Daly, R.I.; Ho, L.; Brookes, J.D. Effect of chlorination on Microcystis aeruginosa cell integrity and subsequent microcystin release and degradation. Environ. Sci. Technol. 2007, 41, 4447–4453. [Google Scholar] [CrossRef]
- Weirich, C.A.; Miller, T.R. Freshwater harmful algal blooms: Toxins and children’s health. Curr Probl Pediatr Adolesc. Health Care 2014, 44, 2–24. [Google Scholar] [CrossRef] [PubMed]
- Dennis, M.A.; Landman, M.; Wood, S.A.; Hamilton, D. Application of flow cytometry for examining phytoplankton succession in two eutrophic lakes. Water Sci. Technol. 2011, 64, 999–1008. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.H.; Lui, D.; Platner, P.J.; Hsu, S.F.; Chu, T.C.; Gaynor, J.J.; Vega, Q.C.; Lustigman, B.K. Induction of temperate cyanophage AS-1 by heavy metal—Copper. BMC Microbiol. 2006, 6, 17. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Chen, W.; Zhang, H.; Peng, L.; Liu, L.; Han, Z.; Wan, N.; Li, L.; Song, L. A flow cytometer based protocol for quantitative analysis of bloom-forming cyanobacteria (Microcystis) in lake sediments. J. Environ. Sci. 2012, 24, 1709–1716. [Google Scholar] [CrossRef]
- Moreira, C.; Ramos, V.; Azevedo, J.; Vasconcelos, V. Methods to detect cyanobacteria and their toxins in the environment. Appl. Microbiol. Biotechnol. 2014, 98, 8073–8082. [Google Scholar] [CrossRef] [PubMed]
- MacColl, R. Cyanobacterial phycobilisomes. J. Struct. Biol. 1998, 124, 311–334. [Google Scholar] [CrossRef] [PubMed]
- Telford, W.G.; Moss, M.W.; Morseman, J.P.; Allnutt, F.C. Cyanobacterial stabilized phycobilisomes as fluorochromes for extracellular antigen detection by flow cytometry. J. Immunol. Methods 2001, 254, 13–30. [Google Scholar] [CrossRef]
- Jin, C.; Mesquita, M.M.F.; Deglint, J.L.; Emelko, M.B.; Wong, A. Quantification of cyanobacterial cells via a novel imaging-driven technique with an integrated fluorescence signature. Sci. Rep. 2018, 8, 9055. [Google Scholar] [CrossRef]
- Van Nevel, S.; Koetzsch, S.; Proctor, C.R.; Besmer, M.D.; Prest, E.I.; Vrouwenvelder, J.S.; Knezev, A.; Boon, N.; Hammes, F. Flow cytometric bacterial cell counts challenge conventional heterotrophic plate counts for routine microbiological drinking water monitoring. Water Res. 2017, 113, 191–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nubel, U.; Garcia-Pichel, F.; Muyzer, G. PCR primers to amplify 16S rRNA genes from cyanobacteria. Appl. Environ. Microbiol. 1997, 63, 3327–3332. [Google Scholar] [Green Version]
- Tillett, D.; Parker, D.L.; Neilan, B.A. Detection of toxigenicity by a probe for the microcystin synthetase A gene (mcyA) of the cyanobacterial genus Microcystis: Comparison of toxicities with 16S rRNA and phycocyanin operon (Phycocyanin Intergenic Spacer) phylogenies. Appl. Environ. Microbiol. 2001, 67, 2810–2818. [Google Scholar] [CrossRef] [PubMed]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, R. Flow Cytometric Analysis for Cyanobacteria in 36 New Jersey Freshwater Bodies. Master’s Thesis, Seton Hall University, South Orange, NJ, USA, 2016. [Google Scholar]
- Paerl, H.W.; Fulton, R.S., 3rd; Moisander, P.H.; Dyble, J. Harmful freshwater algal blooms, with an emphasis on cyanobacteria. Sci. World J. 2001, 1, 76–113. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Otten, T.G. Harmful cyanobacterial blooms: Causes, consequences, and controls. Microb. Ecol. 2013, 65, 995–1010. [Google Scholar] [CrossRef] [PubMed]
- Chu, T.C.; Rienzo, M.J. Bloom-Forming Cyanobacteria and Other Phytoplankton in Northern New Jersey FreshWater Bodies. In International Perspectives on Water Quality Management and Pollutant Control; Quinn, N.W.T., Ed.; IntechOpen: Rijeka, Croatia, 2013; pp. 1–23. [Google Scholar] [CrossRef]
- Cellamare, M.; Rolland, A.; Jacquet, S. Flow cytometry sorting of freshwater phytoplankton. J. Appl. Phycol. 2010, 22, 87–100. [Google Scholar] [CrossRef]
- Poniedzialek, B.; Falfushynska, H.I.; Rzymski, P. Flow cytometry as a valuable tool to study cyanobacteria: A mini-review. Limnol. Rev. 2017, 17, 89–95. [Google Scholar] [CrossRef]
Figure 1.
The 36 water body sites in New Jersey collected from and analyzed in this study. The locations are labeled on the map with the corresponding counties listed.
Figure 1.
The 36 water body sites in New Jersey collected from and analyzed in this study. The locations are labeled on the map with the corresponding counties listed.

Figure 2.
Histograms of phycoerythrin (PE) and allophycocyanin (APC) for three reference cyanobacteria: (A) Synechococcus sp. IU 625; (B) Microcystis aeruginosa; and (C) Cylindrospermum spp. The corresponding fluorescence microscopy images are shown in (A’), (B’) and (C’) respectively.
Figure 2.
Histograms of phycoerythrin (PE) and allophycocyanin (APC) for three reference cyanobacteria: (A) Synechococcus sp. IU 625; (B) Microcystis aeruginosa; and (C) Cylindrospermum spp. The corresponding fluorescence microscopy images are shown in (A’), (B’) and (C’) respectively.

Figure 3.
(A) Sites that contain cyanobacteria including all reference species: Microcystis spp. (orange), Synechococcus sp. IU 625 (blue) and Cylindrospermum spp. (red) related species; (B) Seven sites contained cyanobacteria matched to two reference species: Microcystis spp. (orange) and Synechococcus spp. (blue)-like species; (C) Four sites contained cyanobacteria including Microcystis aeruginosa (orange) and Cylindrospermum spp. (red)-like species; (D) Six sites contained cyanobacteria including Cylindrospermum spp. (red)-like species. (E) Two sites contained cyanobacteria mostly matching to Microcystis spp. (orange)-like species.
Figure 3.
(A) Sites that contain cyanobacteria including all reference species: Microcystis spp. (orange), Synechococcus sp. IU 625 (blue) and Cylindrospermum spp. (red) related species; (B) Seven sites contained cyanobacteria matched to two reference species: Microcystis spp. (orange) and Synechococcus spp. (blue)-like species; (C) Four sites contained cyanobacteria including Microcystis aeruginosa (orange) and Cylindrospermum spp. (red)-like species; (D) Six sites contained cyanobacteria including Cylindrospermum spp. (red)-like species. (E) Two sites contained cyanobacteria mostly matching to Microcystis spp. (orange)-like species.



Figure 4.
Multiple sequence alignment results of 16S rRNA genes from Cylindrospermum spp., Microcystis spp., and Synechococcus spp., by MUSCLE 3.8 [41].
Figure 4.
Multiple sequence alignment results of 16S rRNA genes from Cylindrospermum spp., Microcystis spp., and Synechococcus spp., by MUSCLE 3.8 [41].


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Water properties including temperature, pH and dissolved oxygen of the 36 sites.
| Site Name | Water Temp (°C) | pH | Dissolved O2 (mg/L) |
|---|---|---|---|
| Atlantic City Reservoir | 20.73 | 7.16 | 9.95 |
| NJLM-1286 (Shadow Lake) | 28.49 | 9.18 | 14.64 |
| Deverson Lake | 26.84 | 5.03 | 5.97 |
| Lake Pachoango | 29.35 | 6.79 | 11.96 |
| Lake Pemberton | 28.72 | 6.38 | 10.21 |
| Mt. Misery Lake | 18.41 | 4.09 | 6.35 |
| Mirror Lake | 21.65 | 5.89 | 7.13 |
| NJLM-0315 | 22.35 | 6.24 | 7.05 |
| NJLM-0754 | 22.95 | 6.92 | 9.81 |
| Hands Mill Pond | 28.08 | 4.44 | 7.65 |
| Union Lake | 27.29 | 7.48 | 9.2 |
| Cooper Lake | 24.88 | 9.75 | 19.17 |
| Iona Lake | 24.89 | 6.33 | 5.07 |
| Franklinville Lake | 26.78 | 5.53 | 3.09 |
| McCarthys Lakes | 26.45 | 6.27 | 8.21 |
| NJLM-0489 | 30.09 | 9.38 | 10.77 |
| Amwell Lake | 28.25 | 8.7 | 9.97 |
| Roosevelt County Park Lake | 29.2 | 8.73 | 13.42 |
| NJLM-1034 | 20.32 | 6.95 | 9.87 |
| Cifrese Lake (NJLM-1045) | 28.17 | 9.2 | 10.18 |
| Lake Morski Oko | 22.64 | 7.0 | 6.57 |
| Prospertown Lake | 27.43 | 6.64 | 7.44 |
| Green Turtle Lake | 28.89 | 8.92 | 8.92 |
| Alloway Lake | 30.9 | 8.03 | 9.59 |
| Parvin Lake | 27.8 | 6.86 | 8.17 |
| Rainbow Lake | 28.1 | 6.83 | 7.11 |
| Great Gorge Lake | 26.57 | 9.24 | 8.89 |
| Hunts Pond | 26.67 | 8.25 | 8.95 |
| Laidlaw Pond | 27.01 | 8.43 | 8.1 |
| Mashipacong Pond | 26.56 | 6.2 | 7.04 |
| NJLM-0213 | 16.97 | 6.03 | 1.66 |
| NJLM-0378 | 25.8 | 6.21 | 2.65 |
| Silver Lake | 26.7 | 6.39 | 3.32 |
| Upper East Highland Lake | 26.87 | 8.51 | 7.92 |
| Watchu Pond | 25.17 | 6.83 | 3.69 |
| Deer Park Pond | 28.2 | 7.15 | 6.81 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Patel, R.; de Oliveira, A.; Newby, R., Jr.; Chu, T. Flow Cytometric Analysis of Freshwater Cyanobacteria: A Case Study. Water 2019, 11, 1422. https://doi.org/10.3390/w11071422
AMA Style
Patel R, de Oliveira A, Newby R Jr., Chu T. Flow Cytometric Analysis of Freshwater Cyanobacteria: A Case Study. Water. 2019; 11(7):1422. https://doi.org/10.3390/w11071422
Chicago/Turabian StylePatel, Ruchit, Aline de Oliveira, Robert Newby, Jr., and Tinchun Chu. 2019. "Flow Cytometric Analysis of Freshwater Cyanobacteria: A Case Study" Water 11, no. 7: 1422. https://doi.org/10.3390/w11071422
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.