Natural Attenuation of Mn(II) in Metal Refinery Wastewater: Microbial Community Structure Analysis and Isolation of a New Mn(II)-Oxidizing Bacterium Pseudomonas sp. SK3


Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection and Analysis of On-Site Samples
2.1.1. Water Samples
2.1.2. Mn-Deposit Sample
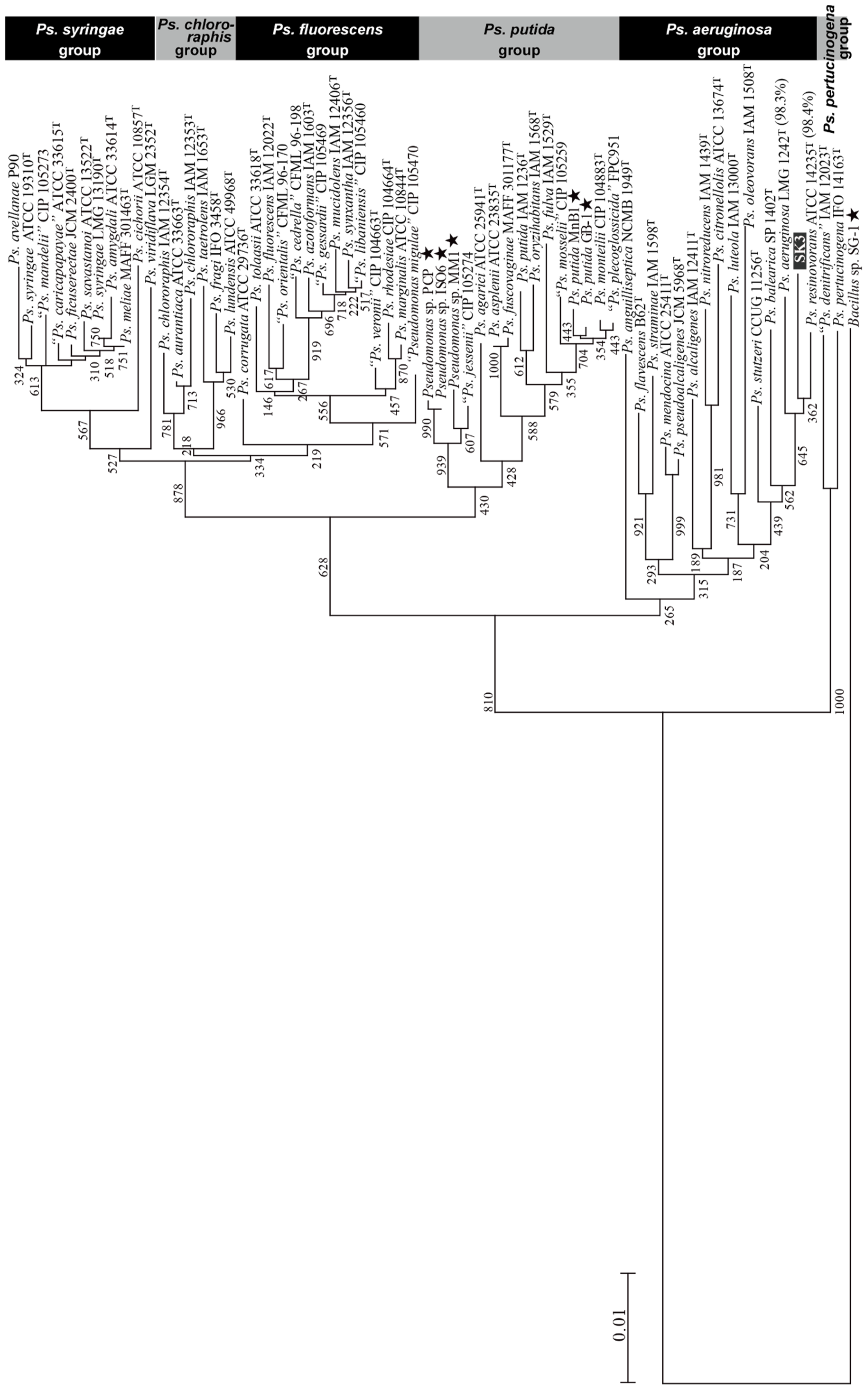
2.2. Isolation, Identification, and Construction of a Phylogenetic Tree for the New Mn(II)-Oxidizing Bacterium
2.3. Mn(II) Oxidation Test
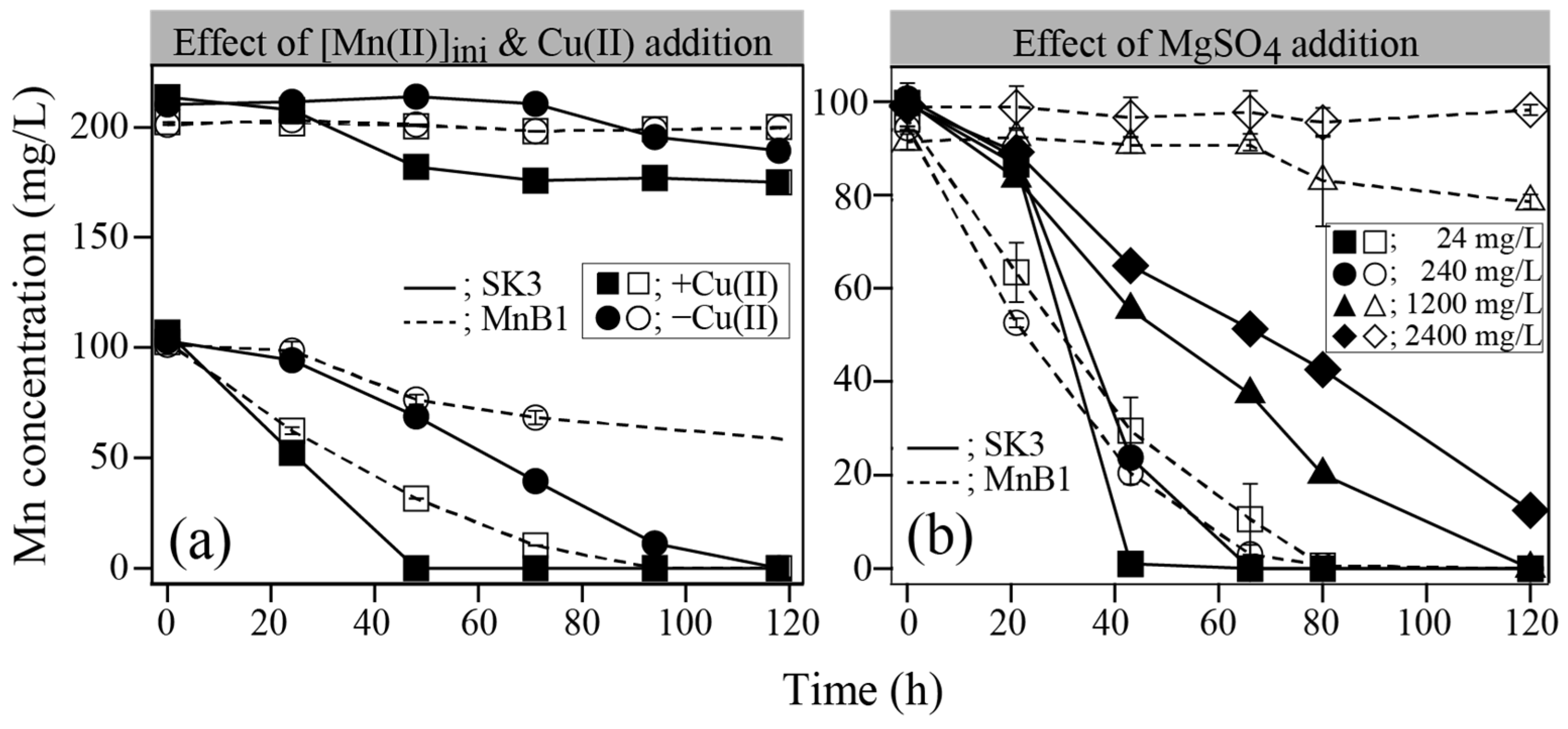
2.3.1. Effect of Initial Mn(II), Cu(II), and MgSO4 Concentrations
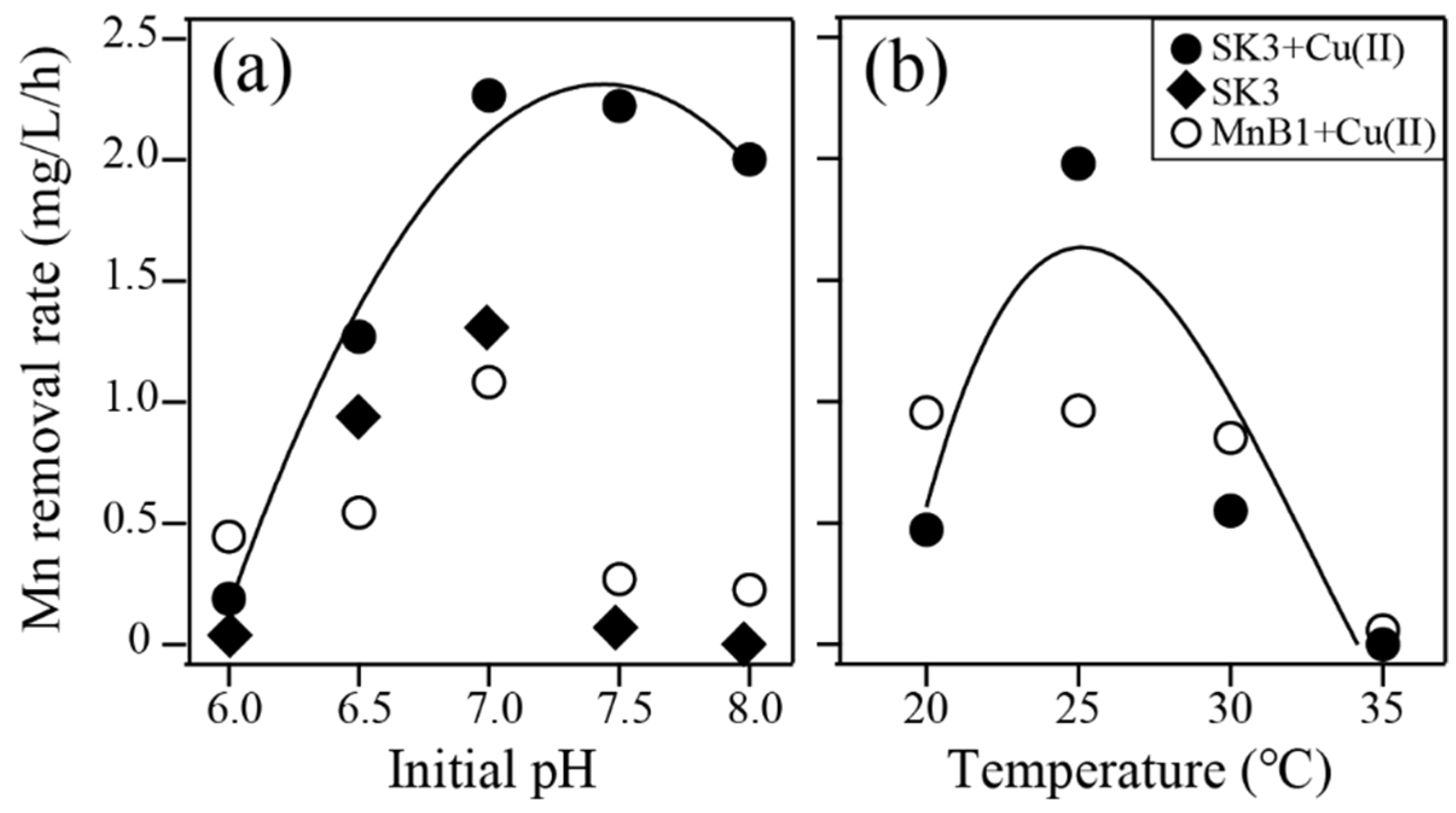
2.3.2. Effect of pH and Temperature
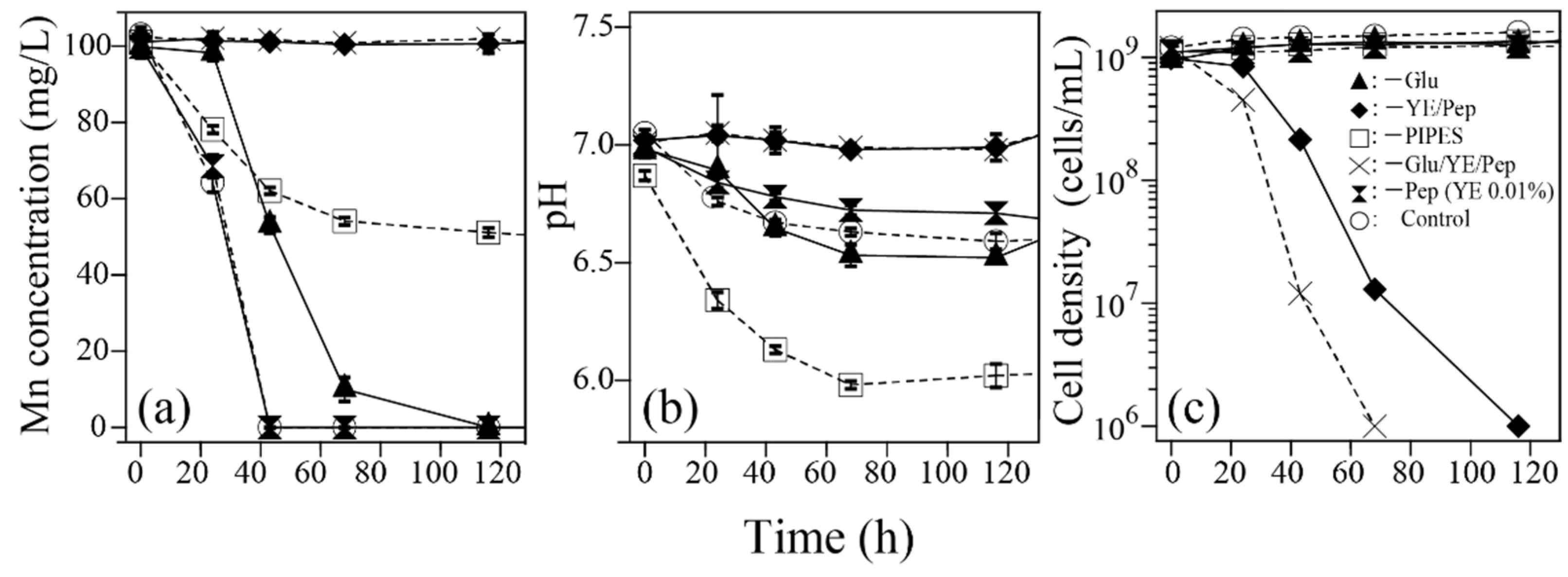
2.3.3. Effect of Individual PYG-1 Medium Components (Test for Isolate SK3 Only)
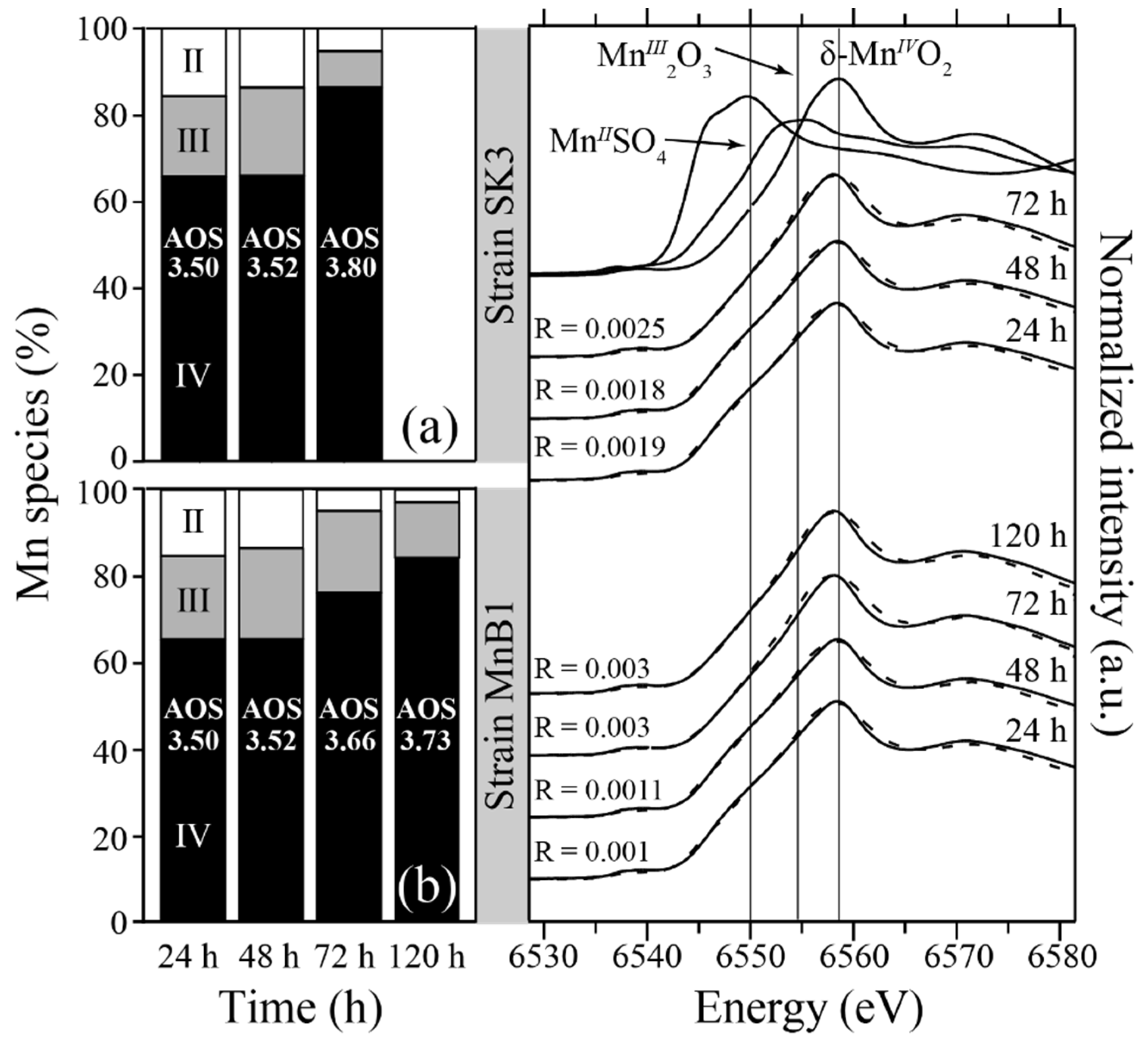
2.4. Characterization of Biogenic Mn-Precipitates
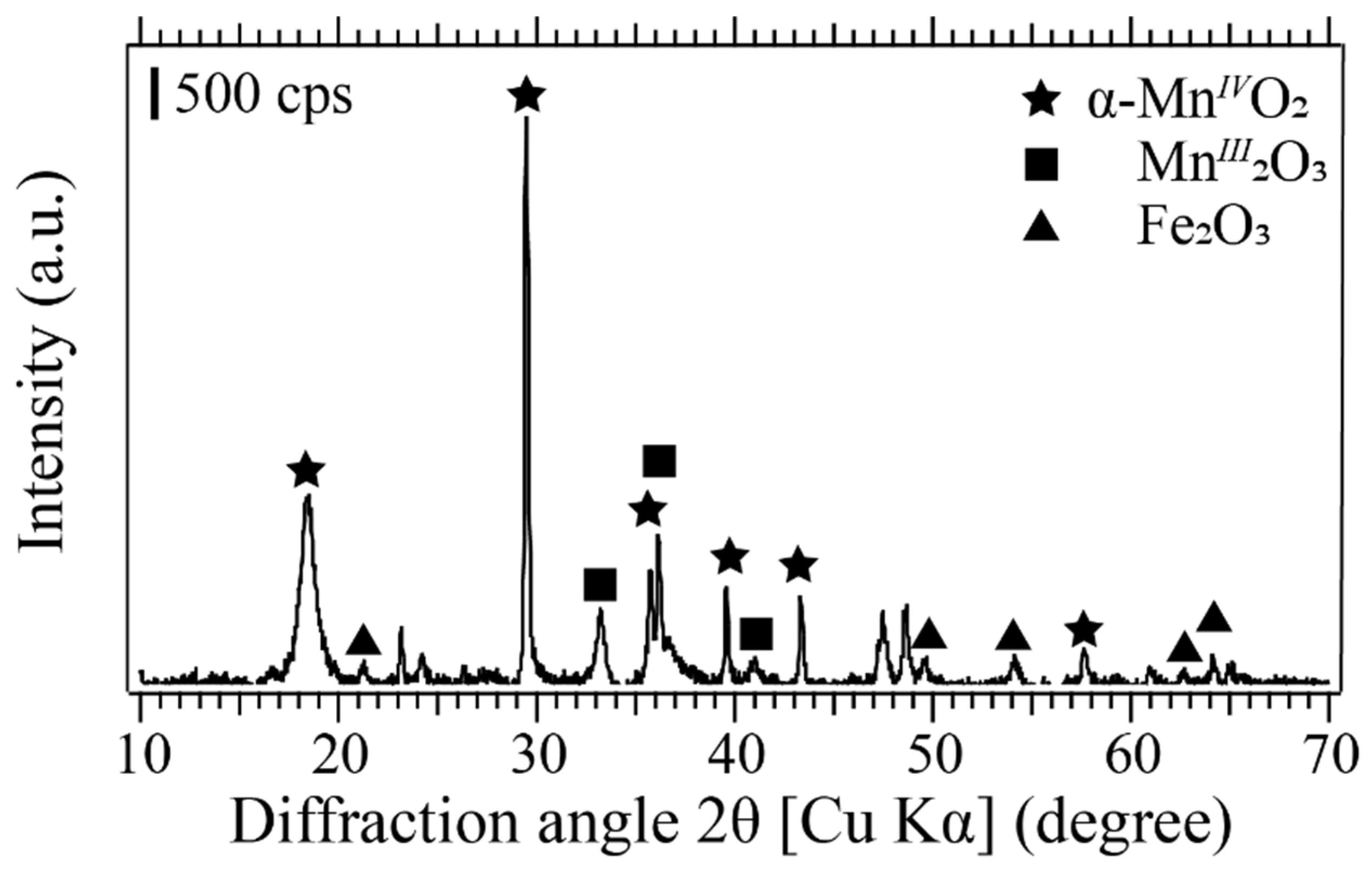
2.4.1. X-ray Diffraction (XRD)
2.4.2. X-ray Absorption Near Edge Structure (XANES)
2.4.3. Scanning Electron Microscope (SEM)
3. Results and Discussion
3.1. Analysis of On-Site Samples
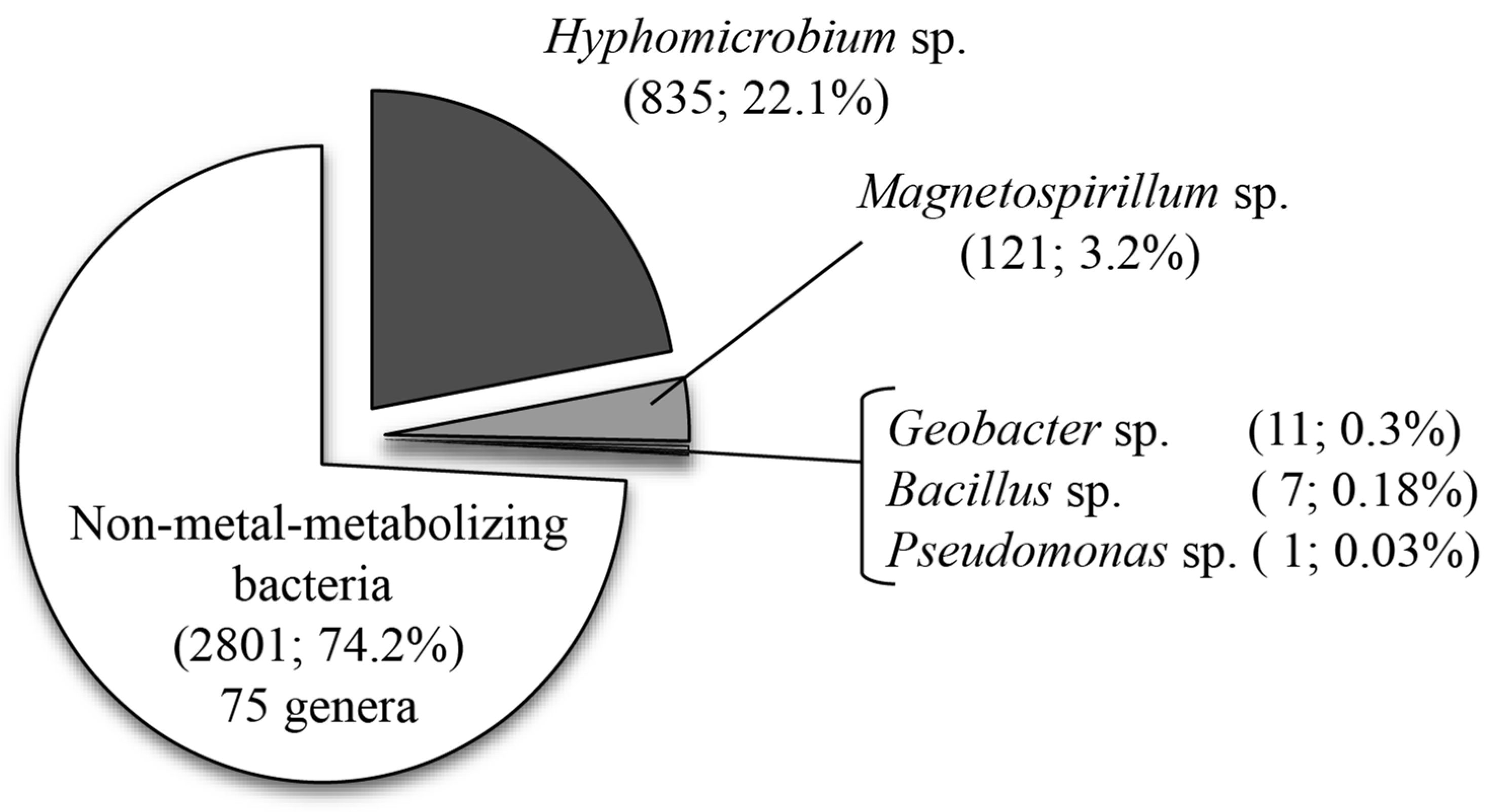
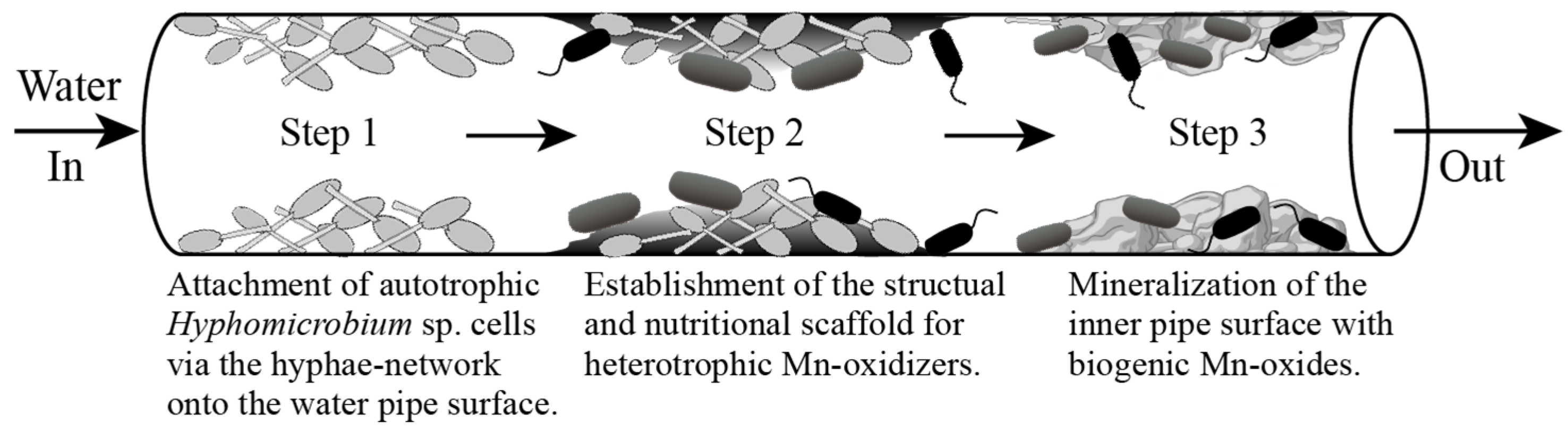
3.2. Microbial Community Structure Analysis
3.3. Isolation of Pseudomonas sp. SK3 and Its Phylogenetic Analysis
3.4. Mn(II) Oxidation by Pseudomonas sp. SK3
3.4.1. Effect of Initial [Mn(II)], [Cu2+], and [MgSO4]
3.4.2. Effect of pH and temperature
3.4.3. Effect of Medium Components
3.4.4. Analysis of Biogenic Mn-Oxides Produced by Pseudomonas sp. SK3
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Morgan, J.J. Kinetics of reaction between O2 and Mn(II) species in aqueous solutions. Geochim. Cosmochim. Acta 2005, 69, 35–48. [Google Scholar] [CrossRef]
- Tebo, B.M.; Bargar, J.R.; Clement, B.G.; Dick, G.J.; Murray, K.J.; Parker, D.; Verity, R.; Webb, S.M. Biogenic manganese oxides: Properties and mechanisms of formation. Annu. Rev. Earth Planet. Sci. 2004, 32, 287–328. [Google Scholar] [CrossRef]
- Tebo, B.M.; Johnson, H.A.; McCarthy, J.K.; Templeton, A.S. Geomicrobiology of manganese(II) oxidation. Trends Microbiol. 2005, 13, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Dick, G.J.; Torpey, J.W.; Beveridge, T.J.; Tebo, B.M. Direct identification of a bacterial manganese(II) oxidase, the multicopper oxidase MnxG, from spores of several different marine Bacillus species. Appl. Environ. Microbiol. 2008, 74, 1527–1534. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Zhang, M.; Liu, Y.; Tang, W. Manganese(II) oxidation by the multi-copper oxidase CopA from Brevibacillus panacihumi MK-8. Enzyme Microb. Technol. 2018, 117, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Corstjens, P.L.A.M.; de Vrind, J.P.M.; Goosen, T.; de Vrind-de Jong, E.W. Identification and molecular analysis of the Leptothrix discophora SS-1 mofA gene, a gene putatively encoding a manganese-oxidizing protein with copper domains. Geomicrobiol. J. 1997, 14, 91–108. [Google Scholar] [CrossRef]
- Francis, C.A.; Tebo, B.M. cumA Multicopper Oxidase Genes from Diverse Mn(II)-oxidizing and non-Mn(II)-oxidizing Pseudomonas Strains. Appl. Environ. Microbiol. 2001, 67, 4272–4278. [Google Scholar] [CrossRef] [PubMed]
- Ridge, J.P.; Lin, M.; Larsen, E.I.; Fegan, M.; McEwan, A.G.; Sly, L.I. A multicopper oxidase is essential for manganese oxidation and laccase-like activity in Pedomicrobium sp. ACM 3067. Environ. Microbiol. 2007, 9, 944–953. [Google Scholar] [CrossRef] [PubMed]
- Geszvain, K.; Smesrud, L.; Tebo, B.M. Identification of a Third Mn(II) Oxidase Enzyme in Pseudomonas putida GB-1. Appl. Environ. Microbiol. 2016, 82, 3774–3782. [Google Scholar] [CrossRef] [PubMed]
- Bohu, T.; Santelli, C.M.; Akob, D.M.; Neu, T.R.; Ciobota, V.; Rosch, P.; Popp, J.; Nietzsche, S.; Kusel, K. Characterization of pH dependent Mn(II) oxidation strategies and formation of a bixbyite-like phase by Mesorhizobium australicum T-G1. Front. Microbiol. 2015, 6, 734. [Google Scholar] [CrossRef] [PubMed]
- Mann, S.; Sparks, N.H.; Scott, G.H.; de Vrind-de Jong, E.W. Oxidation of Manganese and Formation of Mn(3)O(4) (Hausmannite) by Spore Coats of a Marine Bacillus sp. Appl. Environ. Microbiol. 1988, 54, 2140–2143. [Google Scholar] [PubMed]
- Santelli, C.M.; Webb, S.M.; Dohnalkova, A.C.; Hansel, C.M. Diversity of Mn oxides produced by Mn(II)-oxidizing fungi. Geochim. Cosmochim. Acta 2011, 75, 2762–2776. [Google Scholar] [CrossRef]
- Saratovsky, I.; Gurr, S.J.; Hayward, M.A. The Structure of manganese oxide formed by the fungus Acremonium sp. strain KR21-2. Geochim. Cosmochim. Acta 2009, 73, 3291–3300. [Google Scholar] [CrossRef]
- Feng, X.H.; Zhu, M.Q.; Ginder-Vogel, M.; Ni, C.Y.; Parikh, S.J.; Sparks, D.L. Formation of nano-crystalline todorokite from biogenic Mn oxides. Geochim. Cosmochim. Acta 2010, 74, 3232–3245. [Google Scholar] [CrossRef]
- Lefkowitz, J.P.; Rouff, A.A.; Elzinga, E.J. Influence of pH on the reductive transformation of birnessite by aqueous Mn(II). Environ. Sci. Technol. 2013, 47, 10364–10371. [Google Scholar] [CrossRef] [PubMed]
- Okibe, N.; Maki, M.; Sasaki, K.; Hirajima, T. Mn(II)-Oxidizing Activity of Pseudomonas sp. Strain MM1 is Involved in the Formation of Massive Mn Sediments around Sambe Hot Springs in Japan. Mater. Trans. 2013, 54, 2027–2031. [Google Scholar] [CrossRef]
- Holm, N.C.; Gliesche, C.G.; Hirsch, P. Diversity and structure of hyphomicrobium populations in a sewage treatment plant and its adjacent receiving lake. Appl. Environ. Microbiol. 1996, 62, 522–528. [Google Scholar] [PubMed]
- Sly, L.I.; Hodgkinson, M.C.; Arunpairojana, V. Effect of Water Velocity on the Early Development of Manganese-Depositing Biofilm in a Drinking-Water Distribution-System. FEMS Microbiol. Ecol. 1988, 53, 175–186. [Google Scholar] [CrossRef]
- Tyler, P.A. Hyphomicrobia and the oxidation of manganesse in aquatic ecosystems. Antonie Van Leewenhoek 1970, 36, 567–578. [Google Scholar] [CrossRef]
- Cheng, Q.; Nengzi, L.; Bao, L.; Huang, Y.; Liu, S.; Cheng, X.; Li, B.; Zhang, J. Distribution and genetic diversity of microbial populations in the pilot-scale biofilter for simultaneous removal of ammonia, iron and manganese from real groundwater. Chemosphere 2017, 182, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.Y.; Ma, F.; Huang, P.; Yang, J.X. Fe (II) and Mn (II) removal from drilled well water: A case study from a biological treatment unit in Harbin. Desalination 2009, 245, 183–193. [Google Scholar] [CrossRef]
- Bai, Y.; Chang, Y.; Liang, J.; Chen, C.; Qu, J. Treatment of groundwater containing Mn(II), Fe(II), As(III) and Sb(III) by bioaugmented quartz-sand filters. Water Res. 2016, 106, 126–134. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.; Li, D.; Liang, Y.; Zeng, H.; Zhang, J. Operational parameters required for the start-up process of a biofilter to remove Fe, Mn, and NH3-N from low-temperature groundwater. Desalin. Water Treat. 2014, 57, 3588–3596. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X Windows Interface: Flexible Strategies for Multiple Sequence Alignment Aided by Quality Analysis Tools. Nucleic Acids Res. 1998, 25, 4876–4882. [Google Scholar] [CrossRef]
- Villalobos, M.; Toner, B.; Bargar, J.; Sposito, G. Characterization of the manganese oxide produced by Pseudomonas putida strain MnB1. Geochim. Cosmochim. Acta 2003, 67, 2649–2662. [Google Scholar] [CrossRef]
- Ravel, B.; Newville, M. ATHENA, ARTEMIS, HEPHAESTUS: Data analysis for X-ray absorption spectroscopy using IFEFFIT. J. Synchrotron Radiat. 2005, 12, 537–541. [Google Scholar] [CrossRef] [PubMed]
- Friedl, G.; Wehrli, B.; Manceau, A. Solid phases in the cycling of manganese in eutrophic lakes: New insights from EXAFS spectroscopy. Geochim. Cosmochim. Acta 1997, 61, 3277. [Google Scholar] [CrossRef]
- Bodei, S.; Manceau, A.; Geoffroy, N.; Baronnet, A.; Buatier, M. Formation of todorokite from vernadite in Ni-rich hemipelagic sediments. Geochim. Cosmochim. Acta 2007, 71, 5698–5716. [Google Scholar] [CrossRef]
- Bargar, J.R.; Tebo, B.M.; Bergmann, U.; Webb, S.M.; Glatzel, P.; Chiu, V.Q.; Villalobos, M. Biotic and abiotic products of Mn(II) oxidation by spores of the marine Bacillus sp. strain SG-1. Am. Mineral. 2005, 90, 143–154. [Google Scholar] [CrossRef]
- Hiraishi, A.; Yonemitsu, Y.; Matsushita, M.; Shin, Y.K.; Kuraishi, H.; Kawahara, K. Characterization of Porphyrobacter sanguineus sp. nov., an aerobic bacteriochlorophyll-containing bacterium capable of degrading biphenyl and dibenzofuran. Arch. Microbiol. 2002, 178, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Tyler, P.A.; Marshall, K.C. Microbial oxidation of manganese in hydro-electric pipelines. Antonie Van Leewenhoek 1967, 33, 171–183. [Google Scholar] [CrossRef]
- Uebayasi, M.; Tomizuka, N.; Kamibayashi, A.; Tonomura, K. Autotrophic Growth of a Hyphomicrobium sp. and Its Hydrogenase Activity. Agric. Biol. Chem. Tokyo 1981, 45, 1783–1790. [Google Scholar] [CrossRef]
- Zhang, C.; Meng, X.; Li, N.; Wang, W.; Sun, Y.; Jiang, W.; Guan, G.; Li, Y. Two bifunctional enzymes with ferric reduction ability play complementary roles during magnetosome synthesis in Magnetospirillum gryphiswaldense MSR-1. J. Bacteriol. 2013, 195, 876–885. [Google Scholar] [CrossRef] [PubMed]
- Prozorov, T.; Perez Gonzalez, T.; Valverde-Tercedor, C.; Jimenez-Lopez, C.; Yebra-Rodriguez, A.; Körnig, A.; Faivre, D.; Mallapragada, S.; Howse, P.; Bazylinski, D.A.; et al. Manganese Incorporation into Magnetosome Magnetite: Magnetic Signature of Doping. Eur. J. Mineral. 2014, 26, 457–471. [Google Scholar] [CrossRef]
- Zacharoff, L.A.; Morrone, D.J.; Bond, D.R. Geobacter sulfurreducens Extracellular Multiheme Cytochrome PgcA Facilitates Respiration to Fe(III) Oxides but Not Electrodes. Front. Microbiol. 2017, 8, 2481. [Google Scholar] [CrossRef] [PubMed]
- Francis, C.A.; Tebo, B.M. Enzymatic Manganese(II) Oxidation by Metabolically Dormant Spores of Diverse Bacillus Species. Appl. Environ. Microbiol. 2002, 68, 874–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geszvain, K.; McCarthy, J.K.; Tebo, B.M. Elimination of manganese(II,III) oxidation in Pseudomonas putida GB-1 by a double knockout of two putative multicopper oxidase genes. Appl. Environ. Microbiol. 2013, 79, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Solomon, E.I.; Sundaram, U.M.; Machonkin, T.E. Multicopper Oxidases and Oxygenases. Chem. Rev. 1996, 96, 2563–2606. [Google Scholar] [CrossRef] [PubMed]
- Webb, S.M.; Dick, G.J.; Bargar, J.R.; Tebo, B.M. Evidence for the presence of Mn(III) intermediates in the bacterial oxidation of Mn(II). Proc. Natl. Acad. Sci. USA 2005, 102, 5558–5563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, H.; Zhu, M.; Li, W.; Elzinga, E.J.; Villalobos, M.; Liu, F.; Zhang, J.; Feng, X.; Sparks, D.L. Redox Reactions between Mn(II) and Hexagonal Birnessite Change Its Layer Symmetry. Environ. Sci. Technol. 2016, 50, 1750–1758. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wastewater Samples | Mn-Deposits Sample | ||
|---|---|---|---|
| Inlet | Outlet | ||
| pH | 7.9 | 7.6 | - |
| ORP (mV) | 100 | 105 | - |
| TOC (mg/L) | 4 | ND | 2.5 |
| NO3 (mg/L) | 11 | ND | ND |
| Metal Composition | (mg/L) | (mg/g) | |
| Mn | 1–n | 0.n | 617 |
| Fe | < 0.01 | < 0.01 | 121 |
| Cu | < 0.01 | < 0.01 | 0.06 |
| Ca | 389 | 386 | 59 |
| Mg | 280 | 276 | 51 |
| Si | 5.3 | 5.3 | ND |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kitjanukit, S.; Takamatsu, K.; Okibe, N. Natural Attenuation of Mn(II) in Metal Refinery Wastewater: Microbial Community Structure Analysis and Isolation of a New Mn(II)-Oxidizing Bacterium Pseudomonas sp. SK3. Water 2019, 11, 507. https://doi.org/10.3390/w11030507
Kitjanukit S, Takamatsu K, Okibe N. Natural Attenuation of Mn(II) in Metal Refinery Wastewater: Microbial Community Structure Analysis and Isolation of a New Mn(II)-Oxidizing Bacterium Pseudomonas sp. SK3. Water. 2019; 11(3):507. https://doi.org/10.3390/w11030507
Chicago/Turabian StyleKitjanukit, Santisak, Kyohei Takamatsu, and Naoko Okibe. 2019. "Natural Attenuation of Mn(II) in Metal Refinery Wastewater: Microbial Community Structure Analysis and Isolation of a New Mn(II)-Oxidizing Bacterium Pseudomonas sp. SK3" Water 11, no. 3: 507. https://doi.org/10.3390/w11030507