Leaf Gas Exchange, Plant Water Relations and Water Use Efficiency of Vigna Unguiculata L. Walp. Inoculated with Rhizobia under Different Soil Water Regimes


Abstract
:1. Introduction
2. Materials and Methods
2.1. The Growing Conditions and Treatments
2.2. Sampling, Measurements and Analyses
2.3. Statistical Analysis
3. Results
3.1. Soil Water Changes
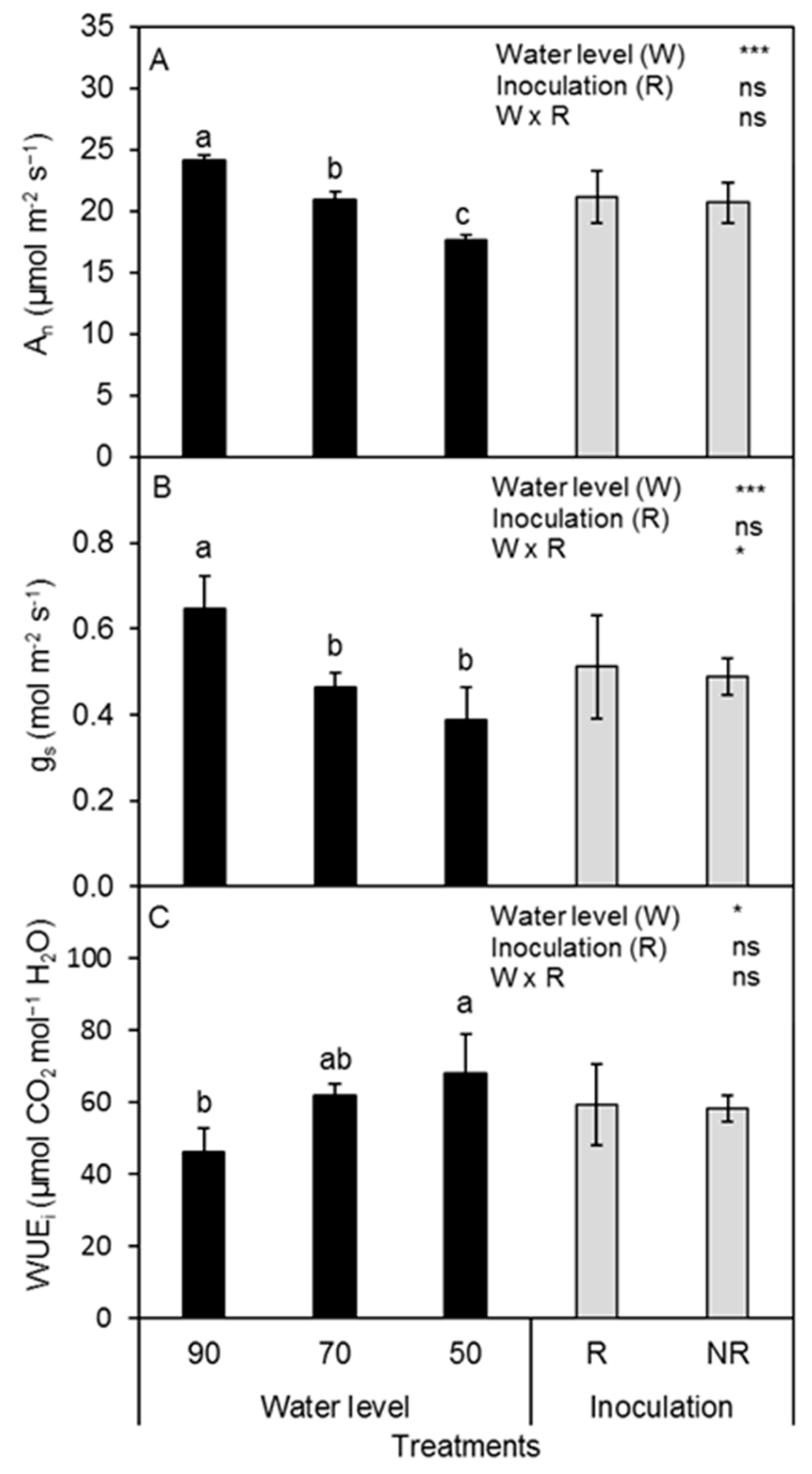
3.2. Leaf Gas Exchange
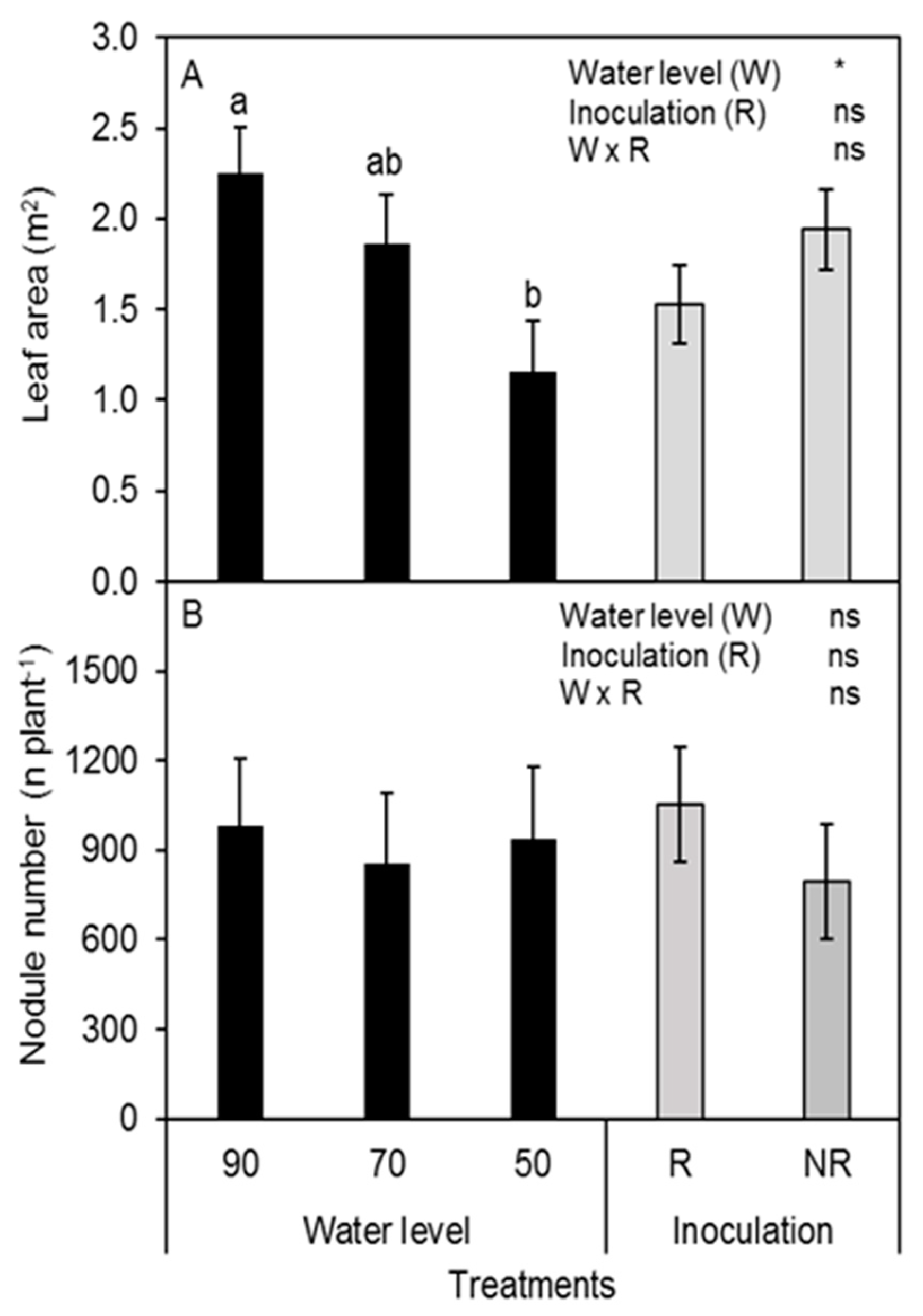
3.3. Leaf Area and Root Nodule Number
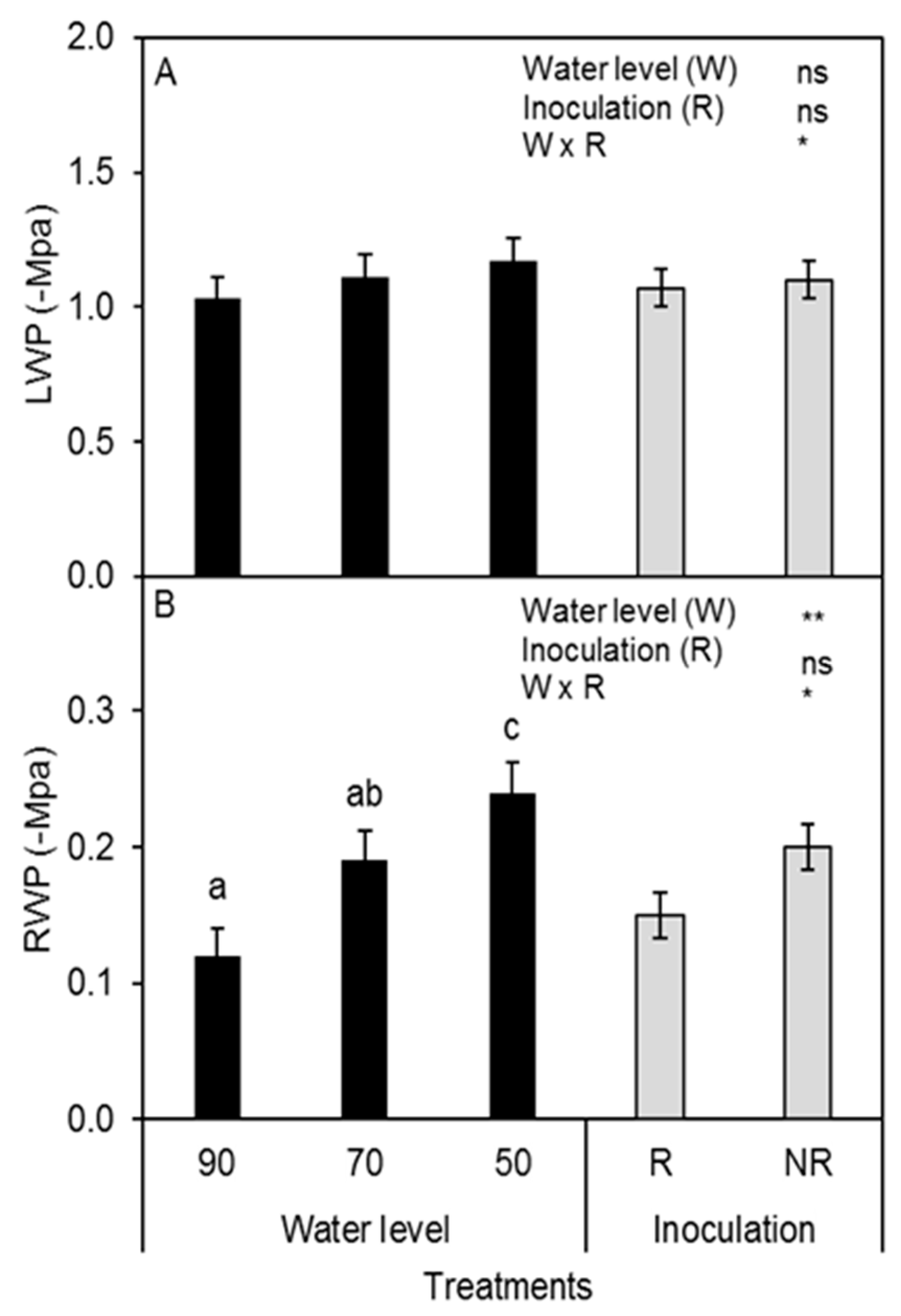
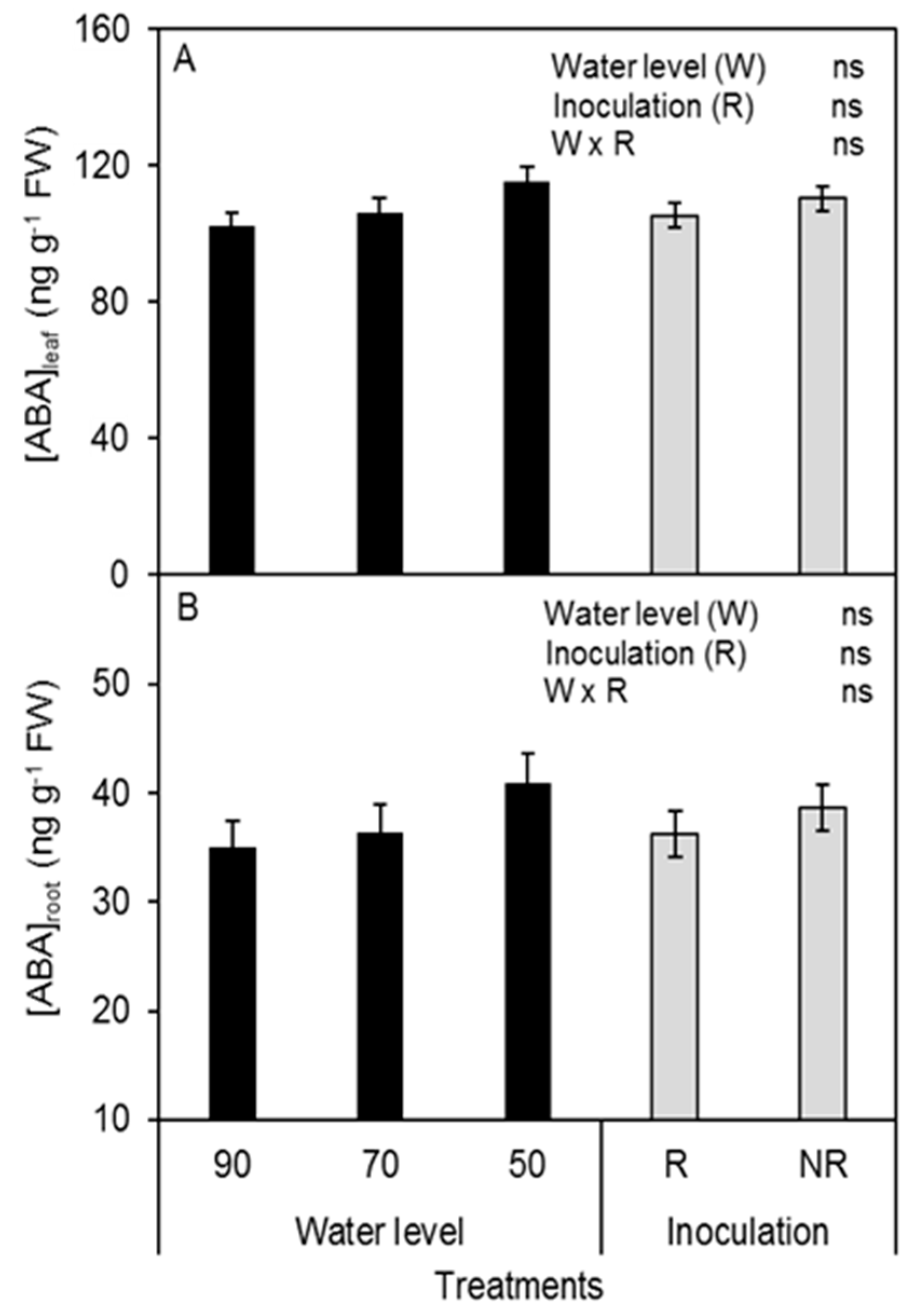
3.4. Plant Water Status and ABA Concentration
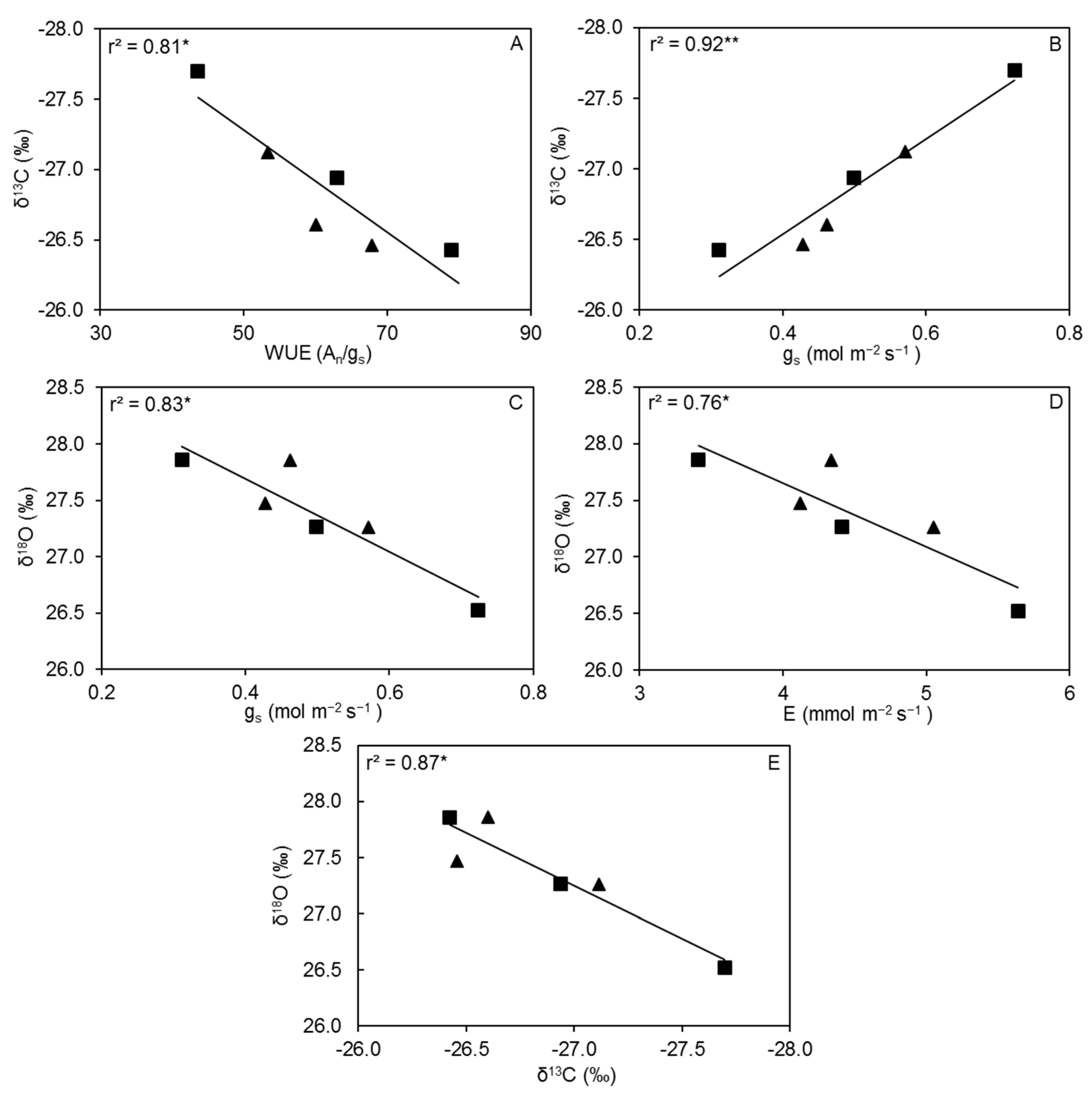
3.5. Biomass Accumulation, Plant Water Use, Water Use Efficiency, Plant δ13C and δ18O
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Donohue, R.J.; Roderick, M.L.; McVicar, T.R.; Farquhar, G.D. Impact of CO2 fertilization on maximum foliage cover across the globe’s warm, arid environments. Geophys. Res. Lett. 2013, 40, 3031–3035. [Google Scholar] [CrossRef]
- Ehlers, J.D.; Hall, A.E. Cowpea (Vigna unguiculata L. Walp.). Field Crops Res. 1997, 53, 187–204. [Google Scholar] [CrossRef]
- Scotti-Camposbr, P.; PhamThibr, A.; Semedobr, J.; Paisbr, I.; Matos, J. Physiological responses and membrane integrity in three Vigna genotypes with contrasting drought tolerance. Emir. J. Food Agric. 2013, 23, 1002. [Google Scholar] [CrossRef]
- Da Silva, D.O.M.; Santos, C.A.F.; Boiteux, L.S. Adaptability and stability parameters of total seed yield and protein content in cowpea (Vigna unguiculata L. Walp.) genotypes subjected to semi-arid conditions. Aust. J. Crop Sci. 2016, 10, 1164–1169. [Google Scholar] [CrossRef]
- Carvalho, M.; Muñoz-amatriaín, M.; Castro, I.; Lino-neto, T.; Matos, M.; Egea-cortines, M.; Rosa, E.; Close, T.; Carnide, V. Genetic diversity and structure of Iberian Peninsula cowpeas compared to world-wide cowpea accessions using high density SNP markers. BMC Genom. 2017, 18, 891. [Google Scholar] [CrossRef] [PubMed]
- Makoi, J.H.J.R.; Chimphango, S.B.M.; Dakora, F.D. Photosynthesis, water-use efficiency and δ13C of five cowpea genotypes grown in mixed culture and at different densities with sorghum. Photosynthetica 2010, 48, 143–155. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon isotope discrimination and photosynthesis. Ann. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Yoo, C.; Pence, H.E.; Hasegawa, P.M.; Mickelbart, M.V. Regulation of transpiration of improving crop water use. Crit. Rev. Plant Sci. 2009, 28, 410–431. [Google Scholar] [CrossRef]
- Anyia, A.O.; Herzog, H. Water-use efficiency, leaf area and leaf gas exchange of cowpeas under mid-season drought. Eur. J. Agron. 2004, 20, 327339. [Google Scholar] [CrossRef]
- Verbree, D.A.; Singh, B.B.; Payne, W.A. Genetics and heritability of shoot drought tolerance in cowpea seedlings. Crop Sci. 2015, 55, 146–153. [Google Scholar] [CrossRef]
- Han, J.M.; Meng, H.F.; Wang, S.Y.; Jiang, C.D.; Liu, F.; Zhang, W.F.; Zhang, Y.L. Variability of mesophyll conductance and its relationship with water use efficiency in cotton leaves under drought pretreatment. J. Plant Physiol. 2016, 194, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Luchi, S.; Kobayashi, M.; Yamaguchi-Shinozaki, K.; Shinozaki, K. A stress-inducible gene for 9-cis-Epoxycarotenoid dioxygenase involved in abscisic acid biosynthesis under water stress in drought-tolerant cowpea. Plant Physiol. 2000, 123, 553–562. [Google Scholar] [CrossRef]
- Luchi, S.; Yamaguchi-Shinozaki, K.; Urao, T.; Terao, T.; Shinozaki, K. Novel drought-inducible genes in the highly drought-tolerant cowpea: Cloning of cDNAs and analysis of their gene expression. Plant Cell Physiol. 1996, 37, 1073–1082. [Google Scholar] [CrossRef]
- Nolan, R.H.; Tarin, T.; Santini, N.S.; McAdam, S.A.M.; Ruman, R.; Eamus, D. Differences in osmotic adjustment, foliar abscisic acid dynamics, and stomatal regulation between an isohydric and anisohydric woody angiosperm during drought. Plant Cell Environ. 2017, 40, 3122–3134. [Google Scholar] [CrossRef]
- Christmann, A.; Weiler, E.W.; Steudle, E.; Grill, E. A hydraulic signal in root-to-shoot signalling of water shortage. Plant J. 2007, 52, 167–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Jensen, C.R.; Shahanzari, A.; Andersen, M.N.; Jacobsen, S.E. ABA regulated stomatal control and photosynthetic water use efficiency of potato (Solanum tuberosum L.) during progressive soil drying. Plant Sci. 2005, 168, 831–836. [Google Scholar] [CrossRef]
- Singh, S.K.; Reddy, R.K. Regulation of photosynthesis, fluorescence, stomatal conductance and water-use efficiency of cowpea (Vigna unguiculata L. Walp.) under drought. J. Photochem. Photobiol. B Biol. 2011, 105, 40–50. [Google Scholar] [CrossRef]
- Munjonji, L.; Ayisi, K.K.; Vandewalle, B.; Haesaert, G.; Boeckx, P. Combining carbon-13 and oxygen-18 to unravel triticale grain yield and physiological response to water stress. Field Crop. Res. 2016, 195, 36–49. [Google Scholar] [CrossRef]
- Pazzagli, P.T.; Weiner, J.; Liu, F. Effects of CO2 elevation and irrigation regimes on leaf gas exchange, plant water relations, and water use efficiency of two tomato cultivars. Agric. Water Manag. 2016, 169, 26–33. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Sharkey, T.D. Stomatal conductance and photosynthesis. Ann. Rev. Plant Physiol. 1982, 33, 317–345. [Google Scholar] [CrossRef]
- Jones, H.G. Plants and Microclimate: A Quantitative Approach to Environmental Plant Physiology, 2nd ed.; Cambridge University Press: Cambridge, UK, 1992. [Google Scholar]
- Wang, Y.; Liu, F.; Andersen, M.N.; Jensen, C.R. Improved plant nitrogen nutrition contributes to higher water use efficiency in tomatoes under alternate partial root-zone irrigation. Funct. Plant Biol. 2010, 37, 175–182. [Google Scholar] [CrossRef]
- Chaves, M.M.; Oliveira, M.M. Mechanisms underlying plant resilience to water deficits: Prospects for water-saving agriculture. J. Exp. Bot. 2004, 55, 2365–2384. [Google Scholar] [CrossRef] [PubMed]
- Herridge, D.F.; Herridge, D. Managing Legume and Fertiliser N for Northern Grains Cropping, 2nd ed.; The GRDC and the University of New England: Armidale, NSW, Australia, 2013. [Google Scholar]
- Serraj, R.; Sinclair, T.R.; Purcell, L.C. Symbiotic N2 fixation response to drought. J. Exp. Bot. 1999, 50, 143–155. [Google Scholar] [CrossRef]
- Jemo, M.; Sulieman, S.; Bekkaoui, F.; Olomide, O.A.K.; Hashem, A.; Abd-Allah, E.F.; Alqarawi, A.A.; Tran, L.P. Comparative Analysis of the Combined Effects of Different Water and Phosphate Levels on Growth and Biological Nitrogen Fixation of Nine Cowpea. Front. Plant Sci. 2017, 8, 2111. [Google Scholar] [CrossRef] [PubMed]
- Yanni, Y.; Zidan, M.; Dazzo, F.; Rizk, R.; Mehesen, A.; Abdelfattah, F.; Elsadany, A. Enhanced symbiotic performance and productivity of drought stressed common bean after inoculation with tolerant native rhizobia in extensive fields. Agric. Ecosyst. Environ. 2016, 232, 119–128. [Google Scholar] [CrossRef]
- Figueiredo, M.V.B.; Burity, H.A.; França, F.P. Water deficit stress effects on N2 fixation in cowpea inoculated with different Bradyrhizobium strains. Can. J. Plant Sci. 1998, 78, 577–582. [Google Scholar] [CrossRef]
- Figueiredo, M.V.; Burity, H.A.; Martínez, C.; Chanway, C.P. Alleviation of drought stress in the common bean (Phaseolus vulgaris L.) by co-inoculation with Paenibacillus polymyxa and Rhizobium tropici. Appl. Soil Ecol. 2008, 40, 182–188. [Google Scholar] [CrossRef]
- Bano, A.; Batool, R.; Dazzo, F. Adaptation of chickpea to desiccation stress is enhanced by symbiotic rhizobia. Symbiosis 2014, 50, 129–133. [Google Scholar] [CrossRef]
- Franzini, V.I.; Azcón, R.; Méndes, F.L.; Aroca, R. Different interaction among Glomus and Rhizobium species on Phaseolus vulgaris and Zea mays plant growth, physiology and symbiotic development under moderate drought stress conditions. Plant Growth Regul. 2013, 70, 265–273. [Google Scholar] [CrossRef]
- Mouradi, M.; Farissi, M.; Bouizgaren, A.; Makoudi, B.; Kabbadi, A.; Very, A.; Sentenac, H.; Qaddourya, A.; Ghoulam, C. Effects of water deficit on growth, nodulation and physiological and biochemical processes in Medicago sativa-rhizobia symbiotic association. Arid Land Res. Manag. 2016, 30, 193–208. [Google Scholar] [CrossRef]
- Giller, K.E. Nitrogen Fixation in Tropical Cropping Systems, 2nd ed.; CABI Publishing: Wallingford, UK, 2001; pp. 253–270. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Richards, R.A. Isotopic Composition of Plant Carbon Correlates with Water-use Efficiency of Wheat Genotypes. Aust. J. Plant Physiol. 1984, 11, 539–552. [Google Scholar] [CrossRef]
- Barbour, M.M. Stable oxygen isotope composition of plant tissue: A review. Funct. Plant Biol. 2007, 34, 83–94. [Google Scholar] [CrossRef]
- Wang, C.; Wu, S.X.; Tankari, M.; Zhang, X.M.; Li, L.; Gong, D.Z.; Hao, W.P.; Zhang, Y.Q.; Mei, X.R.; Wang, Y.F.; et al. Stomatal aperture rather than nitrogen nutrition determined water use efficiency of tomato plants under nitrogen fertigation. Agric. Water Manag. 2018, 209, 94–101. [Google Scholar] [CrossRef]
- Ninou, E.; Tsialtas, J.T.; Dordas, C.A.; Papakosta, D.K. Effect of irrigation on the relationships between leaf gas exchange related traits and yield in dwarf dry bean grown under Mediterranean conditions. Agric. Water Manag. 2013, 116, 235–241. [Google Scholar] [CrossRef]
- Rivas, R.; Falcão, H.M.; Ribeiro, R.V.; Machado, E.C.; Pimentel, C.; Santos, M.G. Drought tolerance in cowpea species is driven by less sensitivity of leaf gas exchange to water deficit and rapid recovery of photosynthesis after rehydration. S. Afr. J. Bot. 2016, 103, 101–107. [Google Scholar] [CrossRef]
- Maia, J.M.; de Macedo, C.E.C.; Voigt, E.L.; Freitas, J.B.S.; Silveira, J.A.G. Antioxidative enzymatic protection in leaves of two contrasting cowpea cultivars under salinity. Biol. Plant. 2010, 54, 159–163. [Google Scholar] [CrossRef]
- Barbosa, M.A.M.; Lobato, A.K.D.S.; Viana, G.D.M.; Coelho, K.N.N.; Barbosa, J.R.S.; Da Costa, R.C.L.; Neto, C.F.D.O. Root contribution to water relations and shoot in two contrasting Vigna unguiculata cultivars subjected to water deficit and inoculation. Rom. Agric. Res. 2012, 30, 155–162. [Google Scholar]
- Davies, W.J.; Wilkinson, S.; Loveys, B. Stomatal control by chemical signalling and the exploitation of this mechanism to increase water use efficiency in agriculture. New Phytol. 2008, 153, 449–460. [Google Scholar] [CrossRef]
- Man, D.; Bao, Y.X.; Han, L.B.; Zhang, X. Drought tolerance associated with proline and hormone metabolism in two tall fescue cultivars. HortScience 2011, 46, 1027–1032. [Google Scholar] [CrossRef]
- Comstock, J.P. Hydraulic and chemical signalling in the control of stomatal conductance and transpriation. J. Exp. Bot. 2002, 53, 195–200. [Google Scholar] [CrossRef]
- Ismail, A.M.; Hall, A.E.; Bray, E.A. Drought and pot size effects on transpiration efficiency and carbon-isotope discrimination of cowpea accessions and hybrids. Aust. J. Plant Physiol. 1994, 21, 23–35. [Google Scholar] [CrossRef]
- Verbree, D.A.; Singh, B.B.; Payne, W.A. Role of gas exchange, leaf water status, and carbohydrate partitioning during the early vegetative stage on drought tolerance in cowpea. Crop Sci. 2014, 54, 770–778. [Google Scholar] [CrossRef]
- Tairo, E.; Mtei, K.; Ndakidemi, P. Influence of Water Stress and Rhizobial Inoculation on the Accumulation of Chlorophyll in Phaseolus vulgaris (L.) Cultivars. Int. J. Plant Soil Sci. 2017, 15, 1–13. [Google Scholar] [CrossRef]
- Ismail, A.M.; Hall, A.E. Correlation between water-use efficiency and carbon isotope discrimination in diverse cowpea genotypes and isogenic lines. Crop Sci. 1992, 32, 7–12. [Google Scholar] [CrossRef]
- Tomás, M.; Medrano, H.; Pou, A.; Escalona, J.M.; Martorell, S.; Ribas-Carbó, M.; Flexas, J. Water-use efficiency in grapevine cultivars grown under controlled conditions: Effects of water stress at the leaf and whole-plant level. Aust. J. Grape Wine Res. 2012, 18, 164–172. [Google Scholar] [CrossRef]
- Flanagan, L.B.; Farquhar, G.D. Variation in the carbon and oxygen isotope composition of plant biomass and its relationship to water-use efficiency at the leaf- and ecosystem-scales in a northern Great Plains grassland. Plant Cell Environ. 2014, 37, 425–438. [Google Scholar] [CrossRef]
- Martin, B.; Thorstenson, Y.R. Stable Carbon Isotope Composition (δ13C), Water Use Efficiency, and Biomass Productivity of Lycopersicon esculentum, Lycopersicon pennellii, and the F1 Hybrid. Plant Physiol. 1988, 88, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, X.; Liu, X.; Zhang, X.; Shao, L.; Sun, H.; Chen, S. The effects of nitrogen supply and water regime on instantaneous WUE, time-integrated WUE and carbon isotope discrimination in winter wheat. Field Crops Res. 2013, 144, 236–244. [Google Scholar] [CrossRef]
- Ellsworth, P.Z.; Ellsworth, P.V.; Cousins, A.B. Relationship of leaf oxygen and carbon isotopic composition with transpiration efficiency in the C4 grasses Setaria viridis and Setaria Italica. J. Exp. Bot. 2017, 68, 3513–3528. [Google Scholar] [CrossRef]
- Condon, A.G.; Richards, R.A.; Rebetzke, G.J.; Farquhar, G.D. Breeding for high water-use efficiency. J. Exp. Bot. 2004, 55, 2447–2460. [Google Scholar] [CrossRef] [Green Version]
- Farquhar, G.D.; Barbour, M.M.; Henry, B.K. Interpretation of oxygen isotope composition of leaf material. In Stable Isotopes: Integration of Biological, Ecological and Geochemical Processes; Griffiths, H., Ed.; Bios Scientific Publishers: Oxoford, UK, 1998; pp. 27–48. [Google Scholar]
- Scheidegger, Y.; Saurer, M.; Bahn, M.; Siegwolf, R. Linking stable oxygen and carbon isotopes with stomatal conductance and photosynthetic capacity: A conceptual model. Oecologia 2000, 125, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Barbour, M.M.; Fischer, R.A.; Sayre, K.D.; Farquhar, G.D. Oxygen isotope ratio of leaf and grain material correlates with stomatal conductance and grain yield in irrigated wheat. Aust. J. Plant Physiol. 2000, 27, 625–637. [Google Scholar] [CrossRef]
- Grams, T.E.E.; Kozovits, A.R.; Häberle, K.H.; Matyssek, R.; Dawson, T.E. Combining δ13C and δ18O analyses to unravel competition, CO2 and O3 effects on the physiological performance of different-aged trees. Plantcell Environ. 2007, 30, 1023–1034. [Google Scholar]
- Roden, J.S.; Farquhar, G.D. A controlled test of the dual-isotope approach for the interpretation of stable carbon and oxygen isotope ratio variation in tree rings. Tree Physiol. 2012, 32, 490–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathu, S.; Herrmann, L.; Pypers, P.; Matiru, V.; Mwirichia, R.; Lesueur, D. Potential of indigenous bradyrhizobia versus commercial inoculants to improve cowpea (Vigna unguiculata L. walp.) and green gram (Vigna radiata L. wilczek.) yields in Kenya. Soil Sci. Plant Nutr. 2012, 58, 750–763. [Google Scholar] [CrossRef]
- Aliyu, I.A.; Yusuf, A.A.; Abaidoo, R.C. Response of grain legumes to rhizobial inoculation in two savanna soils of Nigeria. Afr. J. Microbiol. Res. 2013, 15, 1332–1342. [Google Scholar] [CrossRef]
- Ulzen, J.; Abaidoo, R.C.; Mensah, N.E.; Masso, C.; Abdelgadir, A.H. Bradyrhizobium Inoculants Enhance Grain Yields of Soybean and Cowpea in Northern Ghana. Front. Plant Sci. 2016, 7, 1–9. [Google Scholar] [CrossRef]
- Diallo, A.T.; Samb, P.I.; Roy-Macauley, H. Water status and stomatal behaviour of cowpea, Vigna unguiculata (L.) Walp, plants inoculated with two Glomus species at low soil moisture levels. Eur. J. Soil Biol. 2001, 37, 187–196. [Google Scholar] [CrossRef]
- Sanginga, N.; Dashiell, K.; Okogun, J.A.; Thottappilly, G. Nitrogen fixation and N contribution by promiscuous nodulating soybeans in the southern Guinea savanna of Nigeria. Plant Soil 1997, 195, 257–266. [Google Scholar] [CrossRef]
- Okogun, J.A.; Sanginga, N. Can introduced and indigenous rhizobia strains compete for nodule formation by promiscuous soybean in the moist savanna agroecological zone of Nigeria? Biol. Fertil. Soils 2003, 38, 26–31. [Google Scholar] [CrossRef]
- Figueiredo, M.V.B.; Burity, H.A.; França, F.P. Alleviaion of water stress effects in cowpea by Bradyrhizobium spp. inoculation. Plant Soil 1999, 207, 67–75. [Google Scholar] [CrossRef]
- Catroux, G.; Hartmann, A.; Revellin, C. Trends in rhizobial inoculant production and use. Plant Soil 2001, 230, 21–30. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | Above-Ground Dry Biomass (g plant−1) | Plant Water Use (L plant−1) | WUE (g L−1) | Plant δ13C (‰) | Plant δ18O (‰) |
|---|---|---|---|---|---|
| Water level | |||||
| 90 | 162.9 ± 10.1a | 48.1 ± 2.1a | 3.4 ± 0.1b | −27.41 ± 0.16b | 26.89 ± 0.27b |
| 70 | 138.5 ± 8.2a | 36.4 ± 1.5b | 3.8 ± 0.1a | −26.67 ± 0.15a | 27.38 ± 0.29ab |
| 50 | 80.9 ± 2.6b | 22.7 ± 0.9c | 3.6 ± 0.1ab | −26.50 ± 0.32a | 27.86 ± 0.10a |
| Significance | *** | *** | * | * | * |
| Inoculation | |||||
| R | 123.0 ± 13.0 | 34.8 ± 3.6 | 3.5 ± 0.1 | −27.03 ± 0.23 | 27.21 ± 0.25 |
| NR | 135.0 ± 11.9 | 37.8 ± 3.4 | 3.6 ± 0.1 | −26.74 ± 0.17 | 27.50 ± 0.18 |
| Significance | ns | ns | ns | ns | ns |
| Water level x Inoculation | |||||
| Significance | ns | ns | ns | ns | ns |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tankari, M.; Wang, C.; Zhang, X.; Li, L.; Soothar, R.K.; Ma, H.; Xing, H.; Yan, C.; Zhang, Y.; Liu, F.; et al. Leaf Gas Exchange, Plant Water Relations and Water Use Efficiency of Vigna Unguiculata L. Walp. Inoculated with Rhizobia under Different Soil Water Regimes. Water 2019, 11, 498. https://doi.org/10.3390/w11030498
Tankari M, Wang C, Zhang X, Li L, Soothar RK, Ma H, Xing H, Yan C, Zhang Y, Liu F, et al. Leaf Gas Exchange, Plant Water Relations and Water Use Efficiency of Vigna Unguiculata L. Walp. Inoculated with Rhizobia under Different Soil Water Regimes. Water. 2019; 11(3):498. https://doi.org/10.3390/w11030498
Chicago/Turabian StyleTankari, Moussa, Chao Wang, Ximei Zhang, Li Li, Rajesh Kumar Soothar, Haiyang Ma, Huanli Xing, Changrong Yan, Yanqing Zhang, Fulai Liu, and et al. 2019. "Leaf Gas Exchange, Plant Water Relations and Water Use Efficiency of Vigna Unguiculata L. Walp. Inoculated with Rhizobia under Different Soil Water Regimes" Water 11, no. 3: 498. https://doi.org/10.3390/w11030498